Introduction
Insulin is a key anabolic hormone regulating glucose, lipid, and protein metabolism. Synthesis and secretion occur in pancreatic β cells located within the islets of Langerhans. The islets also contain α cells, which produce glucagon, and δ cells, which secrete somatostatin. The islets of Langerhans constitute the endocrine portion of the pancreas and coordinate hormonal control of metabolic homeostasis. The exocrine pancreas supports digestion through enzyme secretion. Insulin release is tightly regulated by circulating glucose levels, with glucose-stimulated insulin secretion serving as the primary physiological trigger.
At the molecular level, insulin exerts physiologic effects through binding to the insulin receptor, a transmembrane receptor tyrosine kinase (see Image. Insulin Receptor and Paracrine Interaction in Pancreatic Islet Cells). Binding induces receptor autophosphorylation and activation of intracellular signaling pathways, particularly the PI3K–Akt and MAPK cascades. These signaling systems regulate diverse cellular processes, including glucose transport mediated by glucose transporter type 4 (GLUT4), as well as glycogen synthesis, lipogenesis, protein synthesis, and gene expression. These coordinated actions promote energy storage and maintain metabolic homeostasis.[1][2]
In the postprandial state, increased insulin secretion promotes glucose uptake in muscle and adipose tissue, enhances glycogen synthesis, and inhibits hepatic glucose production, thereby reducing blood glucose levels. Glucagon is released during fasting, exercise, or stress to stimulate glycogenolysis and gluconeogenesis, increasing circulating glucose. Coordinated actions of insulin and glucagon are essential for the maintenance of euglycemia through the precise regulation of glucose production, utilization, and storage.
Beyond glucose regulation, insulin plays a critical role in lipid metabolism by promoting lipogenesis and inhibiting lipolysis, as well as in protein metabolism by enhancing amino acid uptake and protein synthesis. Insulin secretion and activity are influenced by multiple physiological and environmental factors, including the stimulation of stress hormones, such as adrenaline and cortisol, sleep patterns, pregnancy, and illness. These factors modulate insulin sensitivity and overall metabolic balance.[3]
Fundamentals
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Fundamentals
Cellular energy homeostasis depends on an interconnected network of metabolic pathways regulating the synthesis, storage, and breakdown of carbohydrates, lipids, and proteins, with insulin serving as a central regulator. Insulin synthesis and secretion occur in pancreatic β cells within the islets of Langerhans. The islets also contain α cells producing glucagon and δ cells secreting somatostatin. The islets of Langerhans form the endocrine pancreas, coordinating hormonal control of metabolism. The exocrine pancreas facilitates digestion through enzyme secretion.
Insulin exerts physiologic effects by binding to the insulin receptor, inducing receptor autophosphorylation and activation of kinase signaling cascades, including the PI3K–Akt and MAPK pathways. These signaling cascades regulate glucose uptake, glycogen synthesis, lipid synthesis, and protein metabolism. In the postprandial state, insulin promotes glucose uptake, glycogenesis, lipogenesis, and protein synthesis, while suppressing hepatic glucose production and lipolysis. During fasting, stress, or exercise, glucagon release stimulates glycogenolysis, gluconeogenesis, and lipolysis to maintain blood glucose levels.[4]
Carbohydrate metabolism includes the following processes:
- Glycolysis: Glucose breakdown for energy production
- Gluconeogenesis: Glucose synthesis from non-carbohydrate precursors
- Glycogenesis: Glycogen synthesis and storage
- Glycogenolysis: Glycogen breakdown and mobilization
Lipid metabolism encompasses the following:
- Lipogenesis: Fatty acid and triglyceride synthesis
- Lipolysis: Triglyceride breakdown for energy utilization
As mentioned, insulin secretion and activity are influenced by the actions of stress hormones, sleep, pregnancy, illness, and environmental factors. These factors modulate insulin sensitivity and overall metabolic balance, ensuring adequate energy supply under varying physiological conditions.[5]
Cellular Level
Insulin exerts its physiological effects through the activation of insulin receptor–dependent signaling pathways in multiple target tissues, including skeletal muscle, adipose tissue, vascular endothelium, cardiac muscle, and hepatic tissue. The downstream effects of insulin are tissue-specific and reflect the distinct metabolic functions of each organ. In skeletal muscle, adipose tissue, and cardiac muscle, insulin stimulates glucose uptake through the promotion of GLUT4 translocation to the plasma membrane, thereby enhancing intracellular glucose utilization. In vascular endothelium and the myocardium, insulin activates endothelial nitric oxide synthase, increasing nitric oxide production, promoting vasodilation, and improving tissue perfusion. In the liver, insulin suppresses hepatic gluconeogenesis and glycogenolysis while promoting glycogenesis and de novo lipogenesis, facilitating postprandial glucose storage.[6]
Insulin actions vary across tissues and metabolic states. In the fed state, elevated circulating insulin levels promote glucose utilization for immediate energy production while directing excess glucose toward anabolic pathways, including glycogen synthesis and lipogenesis. Coordinated processes support overall metabolic homeostasis.
Following a meal, the liver plays a central role in glucose metabolism through the extraction of glucose from the portal circulation. Hepatic glucose undergoes oxidation for adenosine triphosphate (ATP) production or conversion to glycogen via glycogenesis for storage. When glycogen stores approach capacity, excess glucose is metabolized into fatty acids, which are subsequently esterified into triglycerides and exported as very-low-density lipoproteins for storage in adipose tissue. Glucose also serves as a key precursor for biosynthetic pathways, contributing to the synthesis of fatty acids, nucleotides, and amino sugars required for cellular structure and function.[7]
In the central nervous system, glucose serves as the primary energy substrate. However, uptake is largely insulin-independent and mediated by glucose transporter types 1 (GLUT1) and 3 (GLUT3). Neurons oxidize glucose via glycolysis and oxidative phosphorylation to generate ATP and support neurotransmitter synthesis. Hypoglycemia impairs neuronal function, leading to clinical manifestations, such as confusion, dizziness, and lightheadedness.[8]
Red blood cells depend entirely on glucose as the sole energy source, as mitochondria are absent. Glucose metabolism in erythrocytes occurs via anaerobic glycolysis to generate ATP. In skeletal muscle, glucose undergoes glycolysis during exercise or stress to meet energy demands, while at rest, skeletal muscle stores glucose as glycogen via glycogenesis.
Adipose tissue utilizes glucose for ATP production and the generation of glycerol-3-phosphate, required for triglyceride synthesis. Insulin promotes lipid storage through the stimulation of lipogenesis and inhibition of lipolysis. In addition to its effects on carbohydrate and lipid metabolism, insulin enhances protein synthesis and inhibits proteolysis, contributing to overall anabolic balance.
In summary, circulating glucose concentrations increase in the fed state, stimulating pancreatic β cells to secrete insulin. The hormone facilitates glucose uptake in insulin-sensitive tissues and coordinates metabolic pathways that promote glucose utilization and storage, thereby restoring and maintaining euglycemia.
Molecular Level
Insulin is a peptide hormone synthesized in pancreatic β cells as preproinsulin, consisting of a signal peptide, B chain, C-peptide, and A chain. In the rough endoplasmic reticulum, cleavage of the signal peptide forms proinsulin, which undergoes folding and disulfide bond formation. Proinsulin is transported to the Golgi apparatus and packaged into secretory granules, where cleavage at dibasic residues occurs via prohormone convertases (PC1/3 and PC2) and carboxypeptidase E, producing insulin (A and B chains) and C-peptide. Insulin and C-peptide are secreted in equimolar amounts, making C-peptide a useful marker of endogenous insulin production and enabling differentiation between exogenous insulin administration and insulinoma. Insulin is stored in β-cell granules as zinc-coordinated hexamers and released in response to glucose stimulation.[9][10][11]
Function
Roles of Insulin in Metabolism
As mentioned, insulin exerts broad regulatory effects across multiple metabolic pathways. The most physiologically significant roles are outlined below.
Glucose metabolism
Glucose homeostasis is maintained through insulin-mediated glucose uptake and glucose-stimulated insulin secretion. Insulin increases glucose uptake in skeletal muscle and adipose tissue while decreasing hepatic glucose production.
Mechanistically, insulin binds to the α subunit of the insulin receptor and activates β-subunit tyrosine kinase activity. Activation of the receptor stimulates phosphorylation of insulin receptor substrates, resulting in activation of the PI3K–Akt pathway.
Key effects include increased GLUT4 translocation and enhanced glucose uptake in muscle and adipose tissue. GLUT4-mediated uptake is stimulated by insulin and exercise, with exercise representing an insulin-independent mechanism. Additional effects include increased glycogen synthesis and glycolysis, as well as reduced gluconeogenesis.
Glycogen metabolism
Insulin promotes glycogenesis and inhibits glycogenolysis in the liver (see Image. Glycogen Structure and Metabolism). Key enzyme regulation involves protein phosphatase 1. Dephosphorylation results in the activation of glycogen synthase. Dephosphorylation also results in the inactivation of phosphorylase kinase and glycogen phosphorylase. The net effects include increased glycogen synthesis and decreased glycogen breakdown.
Hepatic gene regulation
Insulin regulates hepatic gene expression by inhibiting gluconeogenesis through downregulation of phosphoenolpyruvate carboxykinase, fructose-1,6-bisphosphatase, and glucose-6-phosphatase. Glycolytic activity is increased through the upregulation of glucokinase and pyruvate kinase. Lipogenesis is also promoted through increased expression of fatty acid synthase, acetyl coenzyme A (acetyl-CoA) carboxylase, and pyruvate dehydrogenase.
Lipid metabolism
Insulin promotes fat storage and inhibits fat breakdown. In mechanistic terms, the hormone increases lipogenesis in the liver and adipose tissue. Insulin also increases glucose uptake, providing glycerol-3-phosphate for triglyceride synthesis. Hormone-sensitive lipase is inhibited via dephosphorylation. The net effect includes reduced circulating free fatty acids.
Protein metabolism
Insulin is an anabolic hormone that regulates protein metabolism. Insulin increases amino acid uptake, particularly alanine, arginine, and glutamine. Insulin also enhances protein synthesis in skeletal muscle and the liver. Proteolysis is decreased through the inhibition of the ubiquitin–proteasome pathway.
Effects on Vascular Function and Inflammatory Processes
Insulin exerts vasodilatory and anti-inflammatory effects through actions on the vascular endothelium and immune cells. In the vascular endothelium, insulin increases endothelial nitric oxide synthase activity, leading to increased nitric oxide production and subsequent vasodilation. Insulin also decreases nuclear factor kappa B (NF-κB) activation, resulting in reduced expression of adhesion molecules, including intercellular adhesion molecule 1 and E-selectin. In macrophages, insulin decreases nicotinamide adenine dinucleotide phosphate oxidase activity, leading to reduced production of reactive oxygen species. Insulin also decreases NF-κB signaling, resulting in reduced synthesis of proinflammatory cytokines, including tumor necrosis factor α, interleukins 6 and 8, monocyte chemoattractant protein 1, and matrix metalloproteinases.
Clinical Significance
Chronic hyperglycemia exerts widespread adverse effects on physiological systems and serves as a key contributor to the development of multiple metabolic diseases. Diabetes mellitus, the most prevalent, is defined as a chronic disorder resulting from either insufficient insulin production by the pancreas, as in type 1 diabetes mellitus (T1D), or ineffective utilization of insulin by target tissues, as in type 2 diabetes mellitus (T2D). These 2 types account for the majority of cases.[12][13]
Core Pathophysiology
The exact etiology of diabetes mellitus remains incompletely understood. Several factors correlate with increased disease prevalence, including genetics, lifestyle, and environmental exposures, such as certain viral infections.
Chronic hyperglycemia results from an imbalance between glucose entry into the circulation, including dietary intake and hepatic glucose production, and glucose uptake and utilization by insulin-dependent tissues. Key defects include decreased peripheral glucose uptake in skeletal muscle and adipose tissue, along with increased hepatic glucose production through gluconeogenesis and glycogenolysis.
T1D results from autoimmune destruction of pancreatic β cells, leading to absolute insulin deficiency. The pathogenesis is mediated by T cells and classified as type IV hypersensitivity. Serologic findings include anti-glutamic acid decarboxylase antibodies, anti-insulin antibodies, and anti–islet cell antibodies. Histopathology demonstrates insulitis, characterized by lymphocytic infiltration of pancreatic islets. Clinical features include polyuria, polydipsia, and weight loss, with increased risk of diabetic ketoacidosis (DKA).[14][15][16][17][18][19][20]
T2D is characterized by insulin resistance with relative insulin deficiency (see Image. Insulin Resistance Mechanism). Disease progression begins with insulin resistance in skeletal muscle and adipose tissue, followed by compensatory hyperinsulinemia and eventual β-cell exhaustion with reduced insulin secretion. Risk factors include obesity, particularly visceral adiposity, and genetic predisposition. Histologic findings include islet amyloid deposition composed of amylin. T2D is more commonly associated with hyperosmolar hyperglycemic state (HHS).
Major Complications
Major complications of diabetes mellitus include acute metabolic emergencies and chronic microvascular and macrovascular damage. Acute complications include DKA and HHS. DKA occurs primarily in T1D and results from absolute insulin deficiency, leading to increased lipolysis and ketone body production. The resultant accumulation of ketones produces high–anion gap metabolic acidosis. Clinical features include Kussmaul respirations, fruity breath odor due to acetone, and dehydration. HHS occurs primarily in T2D and is characterized by severe hyperglycemia without significant ketosis due to residual insulin activity sufficient to suppress ketogenesis. Severe dehydration and altered mental status are characteristic findings.
Chronic complications are classified as microvascular and macrovascular. Microvascular complications include diabetic retinopathy, characterized by microaneurysms and neovascularization; diabetic nephropathy, marked by mesangial expansion, glomerular basement membrane thickening, and Kimmelstiel–Wilson nodules; and diabetic neuropathy, typically presenting with a “glove and stocking” distribution of sensory loss. Macrovascular complications result from accelerated atherosclerosis and include coronary artery disease, cerebrovascular disease leading to stroke, and peripheral arterial disease.
Mechanisms of Chronic Complications
The pathogenesis of chronic complications involves multiple interrelated biochemical pathways contributing to tissue injury in chronic hyperglycemia. Advanced glycation end products result from nonenzymatic glycation of proteins, leading to cross-linking of collagen and subsequent thickening of basement membranes. Clinical effects include diabetic nephropathy, diabetic retinopathy, and accelerated atherosclerosis.
The polyol pathway involves the conversion of glucose to sorbitol via aldose reductase, followed by the transformation of sorbitol to fructose via sorbitol dehydrogenase. Several tissues lack sorbitol dehydrogenase activity, leading to sorbitol accumulation. In the lens, sorbitol trapping within the fibers contributes to cataract formation. In Schwann cells, sorbitol buildup plays a role in peripheral neuropathy. In the kidney, sorbitol retention is implicated in structural and functional damage.
Protein kinase C activation is triggered by chronic hyperglycemia and leads to increased vascular endothelial growth factor expression and greater vascular permeability, resulting in microvascular damage. Oxidative stress results from the increased production of reactive oxygen species, leading to endothelial dysfunction.
Clinical Impact
The increasing prevalence of diabetes mellitus is observed globally, driven primarily by rising obesity rates and an aging population. A substantial proportion of cases remain undiagnosed, contributing to delayed diagnosis and treatment. Diabetes mellitus is a major cause of blindness, end-stage kidney failure, and cardiovascular mortality.
Emerging Therapy
Emerging therapy includes stem cell–derived β cells aimed at restoring endogenous insulin production. Current limitations encompass immature insulin secretion, immune rejection, and poor vascular integration.
Media
(Click Image to Enlarge)
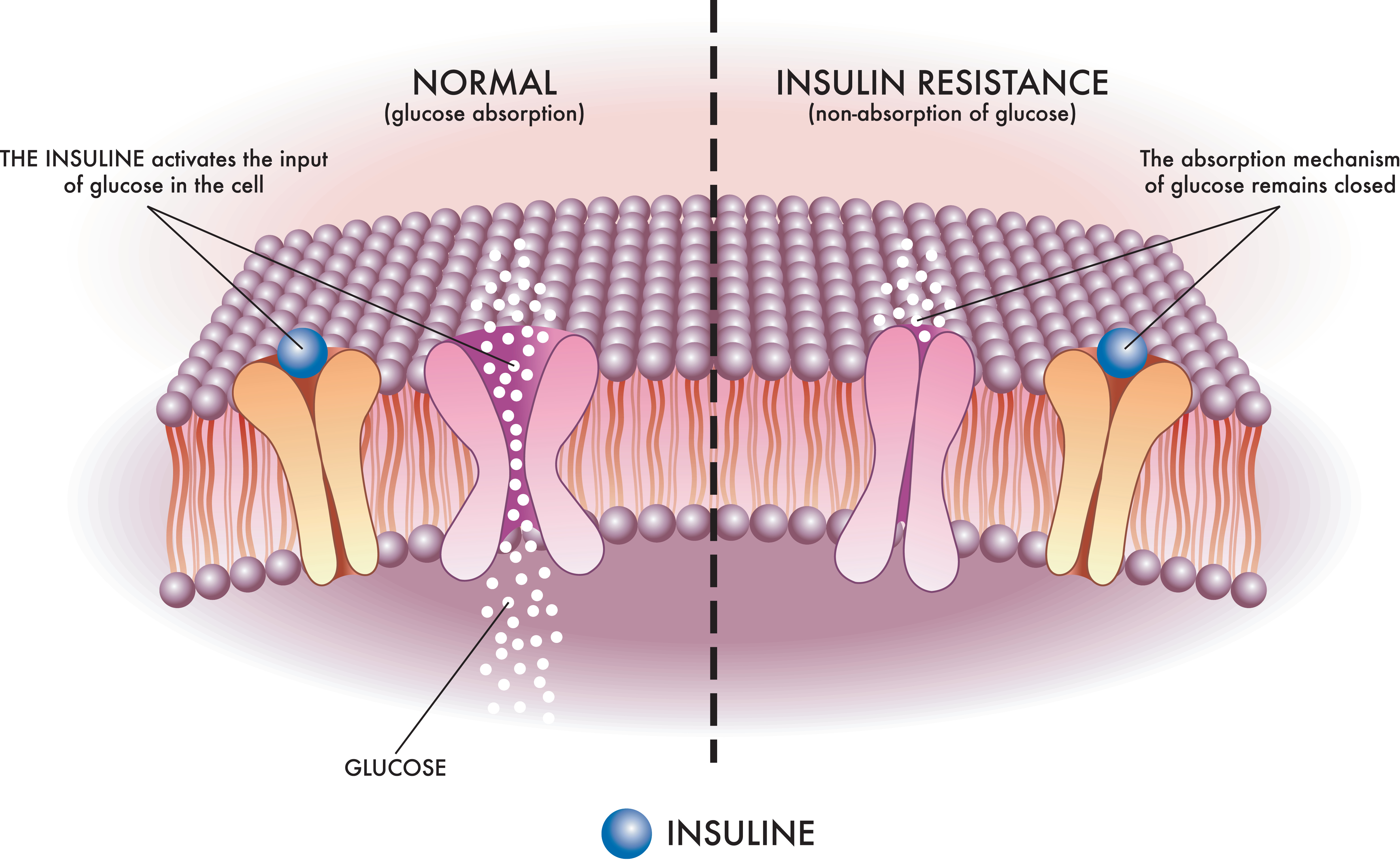
Insulin Resistance Mechanism. This medical illustration depicts the mechanism of insulin resistance, a key contributor to type 2 diabetes, in a clear and schematic manner.
StatPearls - stock.adobe.com
(Click Image to Enlarge)
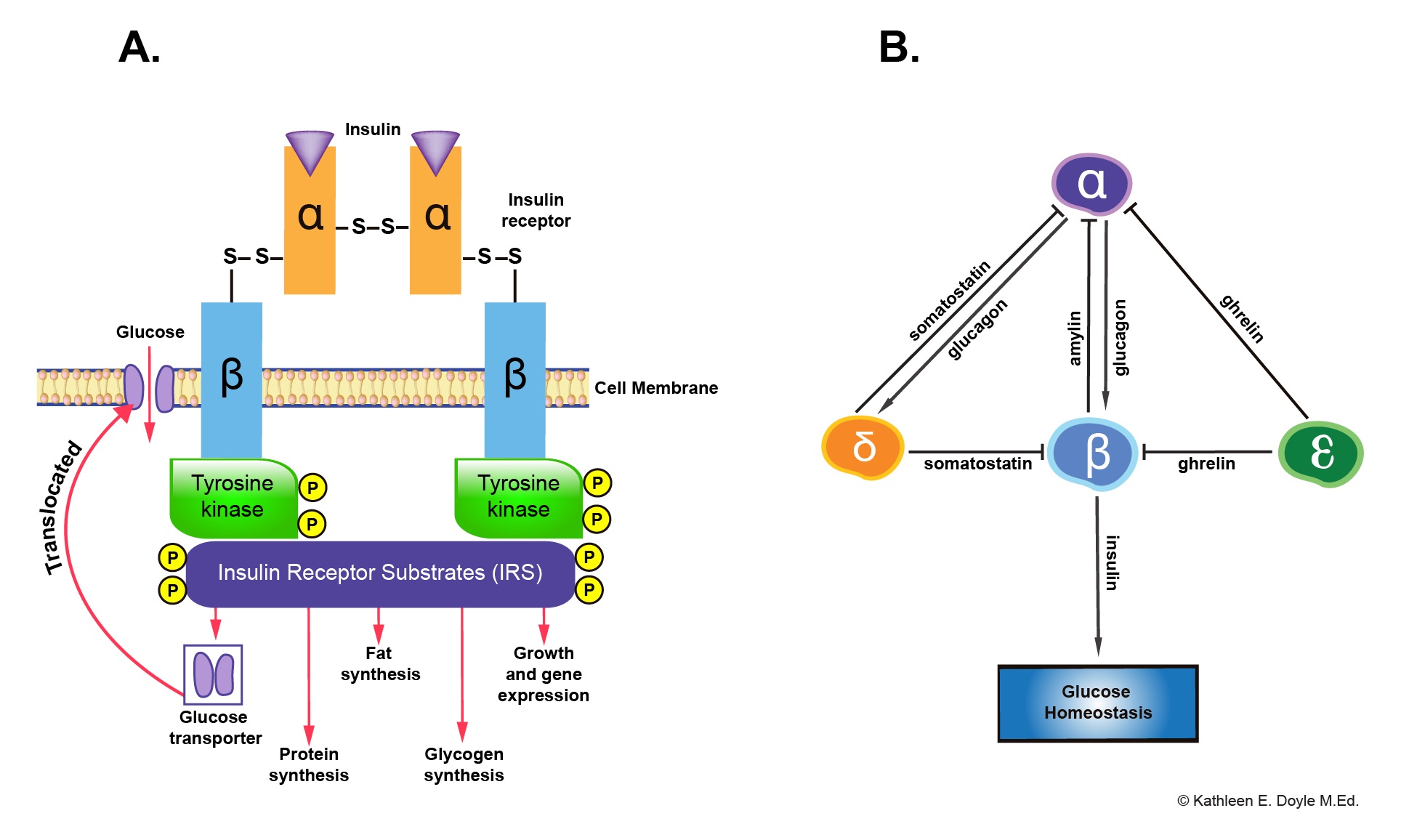
Insulin Receptor and Paracrine Interaction in Pancreatic Islet Cells. Panel A shows the insulin receptor and its downstream functions. Panel B illustrates paracrine interactions among different types of pancreatic islet cells.
Contributed by Kathleen E Doyle MEd, Instructional Designer at the Oakland University William Beaumont School of Medicine.
(Click Image to Enlarge)
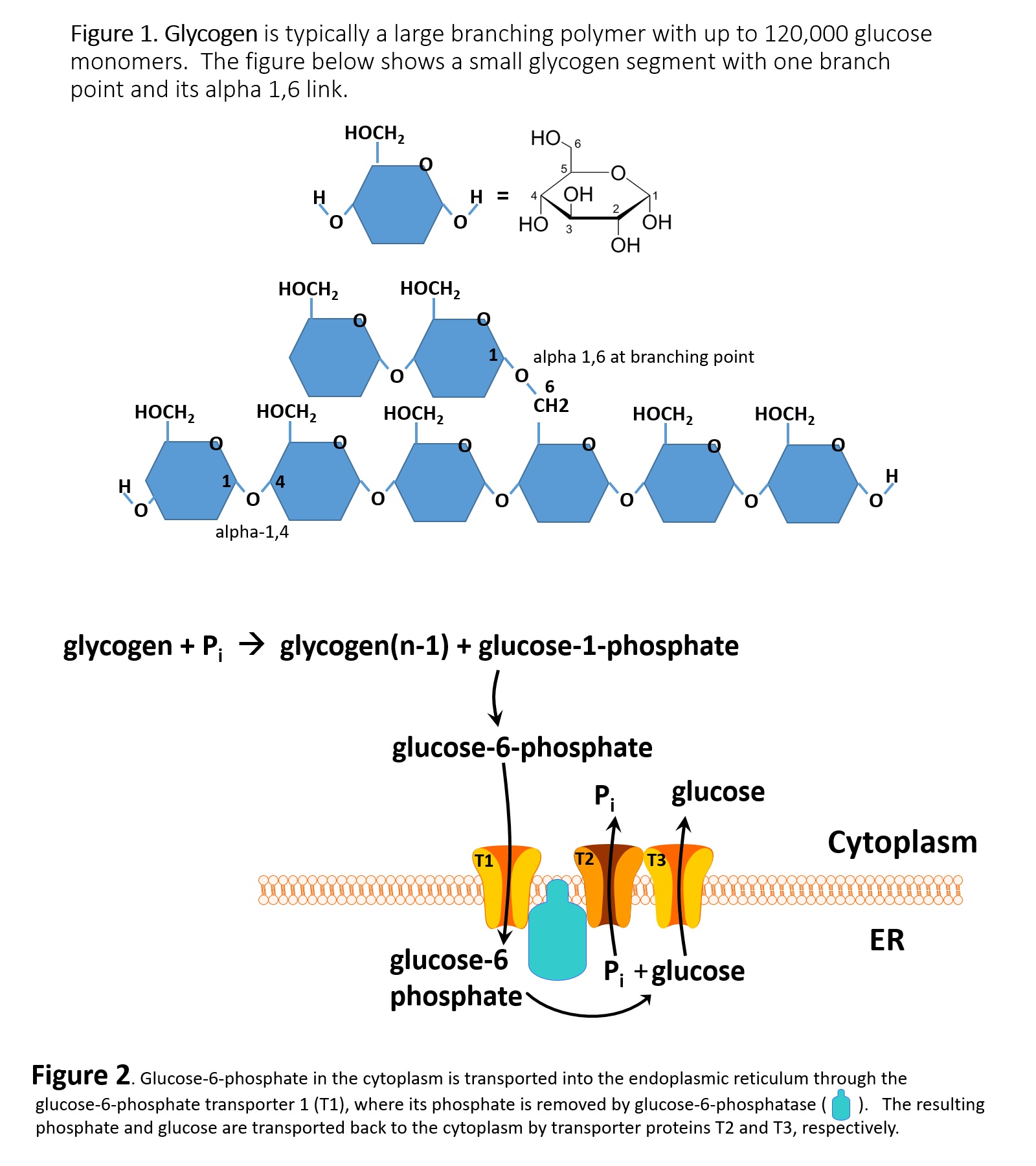
Glycogen Structure and Metabolism. Figure 1 depicts glycogen, a highly branched glucose polymer. Individual glucose monomers form linear chains via α-1,4-glycosidic bonds and branch points via α-1,6-glycosidic bonds, creating a compact structure that can contain up to 120,000 glucose units. Figure 2 illustrates glucose processing and transport. Glycogen phosphorylase releases glucose-1-phosphate, which is converted to glucose-6-phosphate in the cytoplasm. Glucose-6-phosphate enters the endoplasmic reticulum (ER) via the T1 transporter, is dephosphorylated by glucose-6-phosphatase, and the resulting free glucose and phosphate are exported to the cytoplasm through T2 and T3 transporters for release into the bloodstream.
Contributed by Bill Stone, MD
References
Norton L, Shannon C, Gastaldelli A, DeFronzo RA. Insulin: The master regulator of glucose metabolism. Metabolism: clinical and experimental. 2022 Apr:129():155142. doi: 10.1016/j.metabol.2022.155142. Epub 2022 Jan 20 [PubMed PMID: 35066003]
Rahman MS, Hossain KS, Das S, Kundu S, Adegoke EO, Rahman MA, Hannan MA, Uddin MJ, Pang MG. Role of Insulin in Health and Disease: An Update. International journal of molecular sciences. 2021 Jun 15:22(12):. doi: 10.3390/ijms22126403. Epub 2021 Jun 15 [PubMed PMID: 34203830]
Ashcroft FM, Rorsman P. Diabetes mellitus and the β cell: the last ten years. Cell. 2012 Mar 16:148(6):1160-71. doi: 10.1016/j.cell.2012.02.010. Epub [PubMed PMID: 22424227]
Joseph JJ, Golden SH. Cortisol dysregulation: the bidirectional link between stress, depression, and type 2 diabetes mellitus. Annals of the New York Academy of Sciences. 2017 Mar:1391(1):20-34. doi: 10.1111/nyas.13217. Epub 2016 Oct 17 [PubMed PMID: 27750377]
Kusters YH, Barrett EJ. Muscle microvasculature's structural and functional specializations facilitate muscle metabolism. American journal of physiology. Endocrinology and metabolism. 2016 Mar 15:310(6):E379-87. doi: 10.1152/ajpendo.00443.2015. Epub 2015 Dec 29 [PubMed PMID: 26714849]
Verberne AJ, Korim WS, Sabetghadam A, Llewellyn-Smith IJ. Adrenaline: insights into its metabolic roles in hypoglycaemia and diabetes. British journal of pharmacology. 2016 May:173(9):1425-37. doi: 10.1111/bph.13458. Epub 2016 Mar 8 [PubMed PMID: 26896587]
Adeva-Andany MM, Pérez-Felpete N, Fernández-Fernández C, Donapetry-García C, Pazos-García C. Liver glucose metabolism in humans. Bioscience reports. 2016 Dec:36(6):. doi: 10.1042/BSR20160385. Epub 2016 Nov 29 [PubMed PMID: 27707936]
Del Moro L, Rota E, Pirovano E, Rainero I. Migraine, Brain Glucose Metabolism and the "Neuroenergetic" Hypothesis: A Scoping Review. The journal of pain. 2022 Aug:23(8):1294-1317. doi: 10.1016/j.jpain.2022.02.006. Epub 2022 Mar 14 [PubMed PMID: 35296423]
Level 2 (mid-level) evidenceSteiner DF. The proinsulin C-peptide--a multirole model. Experimental diabesity research. 2004 Jan-Mar:5(1):7-14 [PubMed PMID: 15198367]
Orci L, Ravazzola M, Amherdt M, Madsen O, Perrelet A, Vassalli JD, Anderson RG. Conversion of proinsulin to insulin occurs coordinately with acidification of maturing secretory vesicles. The Journal of cell biology. 1986 Dec:103(6 Pt 1):2273-81 [PubMed PMID: 3536964]
Daghlas SA, Rahimi N. Biochemistry, Glycogen. StatPearls. 2026 Jan:(): [PubMed PMID: 30969624]
Lin CL, Wu HC, Yu NC, Liu YC, Chien WC. Factors associated with geriatric syndromes in older adults with type 2 diabetes. Aging clinical and experimental research. 2025 Dec 22:38(1):1. doi: 10.1007/s40520-025-03263-6. Epub 2025 Dec 22 [PubMed PMID: 41427979]
Zhou YC, Liu JM, Zhao ZP, Zhou MG, Ng M. The national and provincial prevalence and non-fatal burdens of diabetes in China from 2005 to 2023 with projections of prevalence to 2050. Military Medical Research. 2025 Jun 2:12(1):28. doi: 10.1186/s40779-025-00615-1. Epub 2025 Jun 2 [PubMed PMID: 40457495]
Rinaldi G, Hijazi A, Haghparast-Bidgoli H. Cost and cost-effectiveness of mHealth interventions for the prevention and control of type 2 diabetes mellitus: a protocol for a systematic review. BMJ open. 2019 Apr 11:9(4):e027490. doi: 10.1136/bmjopen-2018-027490. Epub 2019 Apr 11 [PubMed PMID: 30975686]
Level 1 (high-level) evidenceGiugliano D, De Nicola L, Maiorino MI, Bellastella G, Esposito K. Type 2 diabetes and the kidney: Insights from cardiovascular outcome trials. Diabetes, obesity & metabolism. 2019 Aug:21(8):1790-1800. doi: 10.1111/dom.13743. Epub 2019 May 2 [PubMed PMID: 30969018]
Demircik F, Kirsch V, Ramljak S, Vogg M, Pfützner AH, Pfützner A. Laboratory Evaluation of Linearity, Repeatability, and Hematocrit Interference With an Internet-Enabled Blood Glucose Meter. Journal of diabetes science and technology. 2019 May:13(3):514-521. doi: 10.1177/1932296819841357. Epub 2019 Apr 11 [PubMed PMID: 30974988]
Eleftheriadou I, Tentolouris A, Tentolouris N, Papanas N. Advancing pharmacotherapy for diabetic foot ulcers. Expert opinion on pharmacotherapy. 2019 Jun:20(9):1153-1160. doi: 10.1080/14656566.2019.1598378. Epub 2019 Apr 8 [PubMed PMID: 30958725]
Level 3 (low-level) evidenceKuzulugil D, Papeix G, Luu J, Kerridge RK. Recent advances in diabetes treatments and their perioperative implications. Current opinion in anaesthesiology. 2019 Jun:32(3):398-404. doi: 10.1097/ACO.0000000000000735. Epub [PubMed PMID: 30958402]
Level 3 (low-level) evidenceBalboa D, Barsby T, Lithovius V, Saarimäki-Vire J, Omar-Hmeadi M, Dyachok O, Montaser H, Lund PE, Yang M, Ibrahim H, Näätänen A, Chandra V, Vihinen H, Jokitalo E, Kvist J, Ustinov J, Nieminen AI, Kuuluvainen E, Hietakangas V, Katajisto P, Lau J, Carlsson PO, Barg S, Tengholm A, Otonkoski T. Functional, metabolic and transcriptional maturation of human pancreatic islets derived from stem cells. Nature biotechnology. 2022 Jul:40(7):1042-1055. doi: 10.1038/s41587-022-01219-z. Epub 2022 Mar 3 [PubMed PMID: 35241836]
Hogrebe NJ, Ishahak M, Millman JR. Developments in stem cell-derived islet replacement therapy for treating type 1 diabetes. Cell stem cell. 2023 May 4:30(5):530-548. doi: 10.1016/j.stem.2023.04.002. Epub [PubMed PMID: 37146579]