Introduction
The autonomic nervous system (ANS), a component of the peripheral nervous system, regulates involuntary physiologic processes, including heart rate, blood pressure, respiration, digestion, and sexual arousal. The ANS consists of 3 anatomically distinct divisions: sympathetic, parasympathetic, and enteric (see Image. Autonomic Nervous System Pathways).
The sympathetic (SNS) and parasympathetic (PNS) nervous systems contain afferent fibers that transmit sensory input to the central nervous system (CNS), as well as efferent fibers that convey motor output from the CNS. Motor pathways of the SNS and PNS generally follow a 2-neuron sequence: a preganglionic neuron with a cell body in the CNS and a postganglionic neuron with a cell body in peripheral ganglia that innervates target tissues. The enteric nervous system (ENS) forms an extensive, web-like network capable of autonomous function independent of the CNS.[1] The ENS contains over 100 million neurons of more than 15 morphological types, exceeding the total of all other peripheral ganglia, and primarily regulates digestive processes.[2]
Activation of the SNS induces a state of heightened activity and alertness, commonly referred to as the “fight or flight” response. Physiologic effects include increases in heart rate and blood pressure, stimulation of glycogenolysis, and inhibition of gastrointestinal peristalsis.[3] Sympathetic innervation extends broadly to most organs and tissues, but structures such as cartilage and avascular epithelia lack direct sympathetic input. The PNS facilitates “rest and digest” processes, promoting homeostatic functions, such as modulating heart rate and atrioventricular conduction and restoring gastrointestinal peristalsis and digestion.[4]
Attenuation of arterial pressure primarily occurs indirectly via cardiac output reduction and reflex mechanisms, as systemic vascular tone is predominantly maintained by sympathetic activity. Parasympathetic innervation is largely confined to the head, viscera, and external genitalia, with minimal presence in musculoskeletal tissue and skin, rendering the PNS anatomically smaller than the SNS.[5] The ENS comprises reflex pathways that regulate digestive functions, including smooth muscle contraction and relaxation, secretion and absorption, and local blood flow.[6]
Presynaptic neurons of both the SNS and PNS utilize acetylcholine (ACh) as their neurotransmitter. Postsynaptic sympathetic neurons primarily release norepinephrine to act on target tissues, whereas postsynaptic parasympathetic neurons release ACh exclusively.[7] Enteric neurons employ multiple neurotransmitters, including ACh, nitric oxide, and serotonin.[8]
Structure and Function
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Structure and Function
Sympathetic Nervous System
Sympathetic neurons have cell bodies located in the intermediolateral columns, or lateral horns, of the spinal cord. Presynaptic fibers exit the spinal cord via anterior roots, enter the anterior rami of T1 to L2 spinal nerves, and proceed to the sympathetic trunks through white rami communicantes. Fibers may ascend or descend the sympathetic trunk to reach superior or inferior paravertebral ganglia, respectively, project to adjacent anterior spinal nerve rami via gray rami communicantes, or traverse the trunk without synapsing and continue through abdominopelvic splanchnic nerves to prevertebral ganglia. The central location of sympathetic ganglia results in presynaptic fibers generally being shorter than postsynaptic fibers (see Image. Organization of Sympathetic Efferents).
Paravertebral ganglia form nodular structures along the sympathetic trunk adjacent to the spinal column, where preganglionic and postganglionic neurons synapse. Ganglion numbers vary among individuals, but typically include 3 cervical, 12 thoracic, 4 lumbar, and 5 sacral ganglia. Only the cervical ganglia possess distinct names: superior, middle, and inferior cervical ganglia. The inferior cervical ganglion may fuse with the 1st thoracic ganglion to form the stellate ganglion.[9]
From the paravertebral ganglia, a subset of postganglionic fibers reenters adjacent spinal nerves via gray rami communicantes and subsequently travels in the ventral and dorsal rami to innervate the body wall, including sweat glands, arrector pili muscles, and cutaneous vessels. Visceral branches form the splanchnic nerves: cardiopulmonary splanchnic nerves convey postganglionic fibers to thoracic viscera, whereas abdominopelvic splanchnic nerves (greater, lesser, least, and lumbar) contain preganglionic fibers projecting to prevertebral ganglia. These pathways transmit both afferent and efferent fibers between the CNS and visceral organs (see Image. The Sympathetic Nerves).
Nerves destined for the abdominal and pelvic viscera traverse the paravertebral ganglia without synapsing, forming the abdominopelvic splanchnic nerves. These axonal projections include the greater, lesser, least, and lumbar splanchnic nerves. Presynaptic fibers ultimately synapse in the prevertebral ganglia, located proximal to target organs. Prevertebral ganglia are integral components of nervous plexuses surrounding branches of the aorta, including the celiac, aorticorenal, and superior and inferior mesenteric ganglia.
The greater splanchnic nerve (T5-T9) projects to the celiac ganglion. The lesser splanchnic nerve (T10-T11) innervates the superior mesenteric and aorticorenal or renal ganglia. The least splanchnic nerve (T12) terminates in the renal or aorticorenal ganglion, while the lumbar splanchnic nerves (L1-L2) connect to the inferior mesenteric ganglion.
The celiac ganglion innervates organs derived from the foregut, including the distal esophagus, stomach, proximal duodenum, pancreas, liver, biliary system, spleen, and adrenal glands. The superior mesenteric ganglion supplies derivatives of the midgut, comprising the distal duodenum, jejunum, ileum, cecum, appendix, ascending colon, and proximal transverse colon.
The inferior mesenteric plexus innervates the hindgut, including the distal transverse, descending, and sigmoid colon, rectum, and upper anal canal. Sympathetic supply to pelvic organs, such as the bladder and external genitalia, is transmitted via the hypogastric plexuses, whereas gonadal innervation is mediated by the gonadal plexus, with fibers originating from upper lumbar and lowest thoracic levels.[10][11][12] For more information, see the StatPearls article, Anatomy, Thorax, Greater Splanchnic Nerves.[13]
The 2-neuron rule for SNS and PNS circuits exhibits several exceptions. Postganglionic sympathetic and parasympathetic neurons that synapse onto the ENS functionally form a 3-or-more neuron chain. Preganglionic fibers traverse the celiac plexus without synapsing to innervate adrenal medullary chromaffin cells directly. These cells act as postganglionic effectors, secreting epinephrine directly into the venous system.[14]
Postganglionic sympathetic neurons release norepinephrine, which binds to adrenergic receptors on target tissues. The receptor subtype—α1, α2, β1, β2, or β3—and the tissue distribution of the receptor determine norepinephrine affinity and physiological response.[15] For further details, refer to the StatPearls articles addressing adrenergic receptor structure, function, and signaling.[16][17][18]
As mentioned, the SNS enables the body to react to stressors through the “fight-or-flight” response. This reaction primarily modulates blood vessels, which are tonically innervated. In most cases, an increase in sympathetic signals induces arteriolar vasoconstriction, whereas withdrawal of sympathetic tone permits vasodilation.
Exceptions to this pattern reflect local metabolic control and β2-receptor effects, particularly in skeletal muscle and the coronary circulation. In these regions, the net response during physiologic stress is vasodilation, despite concurrent α-mediated constriction. By contrast, vasodilation of erectile tissue is predominantly mediated by the PNS, with sympathetic activity favoring vasoconstriction and ejaculation. This balance is regulated by α- and β-receptor activity and strong local metabolic influences.
Under physiologic conditions, β-receptor stimulation promotes coronary vessel dilation, but this effect is partially offset by α-receptor-mediated vasoconstriction. In pathologic states, such as coronary artery disease, α-receptor activity is enhanced and β-receptor-mediated vasodilation is attenuated. Consequently, coronary arteries may constrict in response to sympathetic stimulation.[19] Sympathetic activation also increases heart rate and contractile force, which raises metabolic demand and may exacerbate cardiac dysfunction in compromised individuals.[20]
The SNS exhibits tonic activity even in the absence of stressors. In addition to maintaining baseline vascular tone, the SNS contributes to the modulation of airway resistance during the normal respiratory cycle. Airway tone varies with respiration primarily due to phasic vagal activity, which exerts greater bronchoconstrictor influence during expiration, whereas sympathetic bronchodilation provides a background modulatory effect.[21]
The SNS also regulates immune function through innervation of immune organs, including the spleen, thymus, and lymph nodes.[22] Sympathetic input can upregulate or downregulate inflammatory responses.[23] Cells of the adaptive immune system predominantly express β2-receptors, whereas innate immune cells express β2-, α1-, and α2-receptors.[24] Macrophage activity is enhanced by α2-receptor stimulation and suppressed by β2-receptor activation.
The majority of postganglionic sympathetic neurons are noradrenergic and coexpress 1 or more neuropeptides, such as neuropeptide Y or somatostatin. Norepinephrine-neuropeptide Y neurons innervate cardiac blood vessels, thereby regulating myocardial blood flow, whereas norepinephrine-somatostatin neurons of the celiac and superior mesenteric ganglia supply submucosal ganglia of the intestine and contribute to gastrointestinal motility.[25] These neuropeptides are thought to modulate the postsynaptic response to the primary neurotransmitter.
Peptides are also associated with cholinergic sympathetic postganglionic neurons. These neurons predominantly innervate sweat glands and precapillary resistance vessels in skeletal muscle, releasing vasoactive intestinal polypeptide in addition to ACh. Calcitonin gene-related peptide, a potent vasodilator, has also been identified in paravertebral sympathetic neurons.[26][27][28][29]
Parasympathetic Nervous System
Parasympathetic fibers exit the CNS via cranial nerves III (oculomotor), VII (facial), IX (glossopharyngeal), and X (vagus), as well as through the S2 to S4 spinal nerve roots. Four pairs of parasympathetic ganglia are located in the head. Cranial nerve III, through the ciliary ganglion, innervates the iris and ciliary muscles of the eye. Cranial nerve VII innervates the lacrimal, nasal, palatine, and pharyngeal glands via the pterygopalatine ganglion and the sublingual and submandibular glands via the submandibular ganglion. Cranial nerve IX supplies the parotid glands via the otic ganglion.
All other presynaptic parasympathetic fibers synapse in ganglia located near or on the wall of the target tissue, resulting in presynaptic fibers that are significantly longer than the postsynaptic fibers. The term “parasympathetic” historically refers to the craniosacral autonomic outflow that complements the thoracolumbar sympathetic system, with its name originating from the Greek para, meaning “beside.”
The vagus nerve constitutes approximately 75% of the PNS and provides parasympathetic innervation to most thoracic and abdominal viscera. Sacral parasympathetic fibers supply the descending and sigmoid colon as well as the rectum.
Vagal fibers originate from and terminate in medullary nuclei, including the dorsal motor nucleus, which provides parasympathetic output to visceral organs; the nucleus ambiguus, which produces motor fibers and preganglionic neurons that innervate the heart; the nucleus tractus solitarius, which receives visceral and taste afferents; and the spinal trigeminal nucleus, which receives somatic afferent input concerning touch, pain, and temperature from the outer ear, laryngeal mucosa, and portions of the dura.[30][31] The vagus nerve also conveys sensory information from baroreceptors in the aortic arch to the medulla.[32]
As mentioned, the vagus nerve mediates “rest and digest” processes. This cranial nerve promotes cardiac relaxation through several mechanisms. Parasympathetic input decreases contractility in the atria and exerts only minimal influence on the ventricles, reflecting sparse parasympathetic innervation. The vagus nerve primarily reduces conduction velocity through the atrioventricular node. This mechanism underlies the efficacy of carotid sinus massage in terminating atrioventricular nodal reentrant tachycardia and some cases of orthodromic atrioventricular reentrant tachycardia. However, this intervention is not a treatment for Wolff-Parkinson-White syndrome per se, particularly in preexcited atrial fibrillation.
The PNS also exerts a central role in digestion. Parasympathetic fibers to the head stimulate salivation, whereas fibers synapsing onto the ENS enhance peristaltic and secretory activity.[33]
Additionally, the vagus nerve modulates the respiratory cycle. In nonpathological states, parasympathetic activity during expiration contracts and stiffens airways, preventing collapse. This function implicates the PNS in the pathogenesis of postoperative acute respiratory distress syndrome.
The extensive distribution of the vagus nerve has led to its characterization as an “early warning system” for detecting foreign invaders and monitoring systemic recovery. Approximately 80% of vagal fibers are sensory and innervate nearly all major organs. Parasympathetic ganglia express receptors for interleukin 1, a key cytokine in the inflammatory immune response.[34] Activation of these receptors stimulates the hypothalamic-pituitary-adrenal axis and the SNS, resulting in the release of glucocorticoids and norepinephrine, respectively. Vagal activity has been shown to attenuate inflammation via the cholinergic anti-inflammatory pathway. Conversely, vagotomy or administration of anticholinergic agents may exacerbate allergic, asthmatic, or inflammatory responses, whereas vagal stimulation exerts anti-inflammatory effects.
Postganglionic parasympathetic ACh acts on muscarinic receptors (M1, M2, and M3) at effector organs. Nicotinic receptors (N1 and N2) mediate transmission from preganglionic to postganglionic neurons. Postganglionic nicotinic receptors and those on the adrenal medulla are N-type, whereas muscarinic receptors on parasympathetic effectors and sweat glands are M-type. Similar to sympathetic neurons, parasympathetic neurons express and release several neuropeptides, including vasoactive intestinal peptide, neuropeptide Y, and calcitonin gene-related peptide (see Image. Neurotransmitters of the Autonomic Nervous System).[35][36] For additional details on cholinergic receptors, refer to the relevant StatPearls article.[37]
Enteric Nervous System
The ENS consists of 2 ganglionated plexuses: myenteric (Auerbach) and submucosal (Meissner). The myenteric plexus is situated between the longitudinal and circular smooth muscle layers of the gastrointestinal tract, whereas the submucosal plexus resides within the submucosa. The ENS operates as a self-contained system, functioning through local reflex circuits, but frequently receives input from, and provides feedback to, the SNS and PNS. Input may originate from postganglionic sympathetic neurons or preganglionic parasympathetic neurons.[38]
The submucosal plexus regulates the movement of water and electrolytes across the intestinal wall. The myenteric plexus regulates the contraction of the circular and longitudinal muscle layers to produce peristaltic movements.
Motility within the ENS is generated through reflex circuits involving the circular and longitudinal smooth muscle layers. Nicotinic synapses between interneurons mediate these reflex pathways. Upon activation by the presence of a bolus, excitatory neurons in the circular muscle and inhibitory neurons in the longitudinal muscle fire, producing a constricted segment of bowel proximal to the bolus, termed the propulsive segment. Concurrently, excitatory neurons in the longitudinal muscle and inhibitory neurons in the circular muscle fire, creating the receiving segment, into which the bolus progresses. This sequence repeats along successive segments of the bowel.[39]
The ENS exhibits several structural and functional similarities to the CNS. Enteric neurons may be bipolar, pseudounipolar, or multipolar, and neuromodulation occurs via excitatory and inhibitory communication. Furthermore, ENS neurons utilize more than 30 neurotransmitters, many of which are shared with the CNS, with cholinergic and nitrergic transmitters being the most prevalent.[40]
Although much of the discussion has emphasized efferent functions of the ANS, afferent fibers mediate numerous reflex activities that regulate processes ranging from heart rate to immune function. ANS feedback is predominantly processed at a subconscious level, generating reflex responses in visceral or somatic structures. Conscious perception of visceral activity is typically experienced as diffuse pain or cramping, which may correspond to states such as hunger, fullness, or nausea. These sensations most frequently arise from sudden distention or contraction, exposure to chemical irritants, or pathological conditions, including ischemia.[41]
Embryology
Most peripheral neurons and glia, including dorsal root ganglia, autonomic ganglia, and Schwann and satellite cells, derive from the neural crest, whereas somatic motor neurons arise from the neural tube. The neural crest is classically divided into cranial (including cardiac), trunk, vagal, and sacral populations. Dorsal root ganglia and the sympathetic chain, including the adrenal medulla, originate from trunk neural crest cells. Parasympathetic innervation of the heart originates from neural crest-derived Schwann cell precursors of the cardiac-vagal crest lineage.[42] Cranial parasympathetic ganglia also arise from Schwann cell precursors that are neural crest-derived rather than from placodal ectoderm.[43]
The ENS originates from the vagal neural crest, with cells migrating in a rostral-to-caudal pattern through the intestinal wall to form a network of glia and neurons of multiple subtypes.[44] In humans, complete migration and establishment of neural crest cells in the colon occurs by gestational weeks 12 to 13, with neurotransmitter expression and circuit maturation continuing into late gestation and early postnatal life. Mature gut motility is not established until at least late gestation to shortly after birth.[45]
Surgical Considerations
Horner syndrome is an uncommon condition that typically presents with unilateral ptosis, miosis with dilation lag and preserved light reactivity, and facial anhidrosis, which is more often observed in central or preganglionic lesions than in postganglionic lesions. These manifestations result from the disruption of sympathetic fibers within the oculosympathetic pathway. Common etiologies include carotid artery dissection, apical lung (Pancoast) tumors, lateral medullary infarction, and iatrogenic injury from neck or thoracic surgery, including thyroidectomy.[46] Central (1st-order) lesions generally produce Horner syndrome accompanied by brainstem signs such as vertigo, ataxia, and sensory deficits.[47] For additional details, refer to the associated StatPearls articles.[48][49]
Hyperhidrosis is a common disorder characterized by excessive sweating, most frequently affecting the face, palms, soles, and axillae. The etiology of primary hyperhidrosis is not fully understood but is thought to involve increased sympathetic cholinergic stimulation of eccrine sweat glands. Management may be clinical or surgical.[50]
First-line treatment includes topical aluminum chloride, tap-water iontophoresis for the palms and soles, and botulinum toxin A injections for the axillae, palms, or face. Oral anticholinergics, such as oxybutynin and glycopyrrolate, may be employed for refractory cases. Other agents, including clonidine, calcium channel blockers, and gabapentin, have limited supporting evidence.[51]
The most common and definitive surgical intervention is endoscopic thoracic sympathectomy, which may involve resection or ablation. Nerve clipping is potentially reversible if performed early. Although the procedure is permanent, it may produce compensatory hyperhidrosis in a small subset of patients. These symptoms may be equivalent to or more severe than the preoperative state, likely due to mechanisms that remain incompletely understood. Limited evidence suggests that surgical reconstruction of the sympathetic chain can attenuate this compensatory response.[52]
Clinical Significance
The extensive distribution of the ANS renders it susceptible to a wide range of inherited and acquired conditions, including the following:
- Inherited
- Variant (hereditary) transthyretin amyloidosis
- Fabry disease
- Hereditary sensory autonomic neuropathy
- Porphyrias
- Acquired
- Diabetes mellitus
- Immunoglobulin light-chain (AL) and serum amyloid A (AA) amyloidosis
- Uremic neuropathy and chronic liver disease
- Vitamin B12 deficiency
- Toxin- or drug-induced neuropathies, such as from alcohol, amiodarone, or chemotherapy
- Infectious etiologies, including botulism, Chagas disease, HIV, leprosy, Lyme disease, and tetanus
- Autoimmune disorders, including Guillain-Barré syndrome, Lambert-Eaton myasthenic syndrome, rheumatoid arthritis, Sjögren syndrome, and systemic lupus erythematosus
- Neurological disorders, such as multiple system atrophy (Shy-Drager syndrome), Parkinson disease, and Lewy body dementia
- Conditions associated with neoplasia, such as brain tumors and paraneoplastic syndromes [53][54]
Autonomic neuropathy can affect multiple organ systems, producing a broad spectrum of clinical manifestations. Orthostatic hypotension is the most common feature of autonomic dysautonomia, although numerous other findings may also be present.
Cardiovascular manifestations include a fixed heart rate, postural hypotension, and resting tachycardia. Gastrointestinal involvement may present as dysphagia, gastroparesis with associated nausea, vomiting, and abdominal fullness, and constipation. Genitourinary dysfunction can manifest as bladder atony. Pupillary abnormalities may include absent or delayed light reflexes and decreased pupil size. Sexual dysfunction can present as erectile dysfunction in male persons or retrograde ejaculation. Sudomotor involvement may lead to anhidrosis or gustatory sweating. Vasomotor disturbances include cold extremities due to impaired vasoconstrictor responses and dependent edema from loss of venous tone.
Symptoms of orthostatic hypotension most commonly include lightheadedness, tunnel vision, and discomfort in the head, neck, or chest. In some patients, orthostatic hypotension may coexist with supine hypertension caused by increased peripheral resistance, which induces natriuresis and further exacerbates postural hypotension. Additional physiologic stimuli can transiently influence blood pressure. Standing, food ingestion, Valsalva maneuver, dehydration, exercise, and hyperventilation may decrease blood pressure, whereas lying supine, water ingestion, caffeine intake, head-down tilt, and hypoventilation may increase this parameter.
Evaluation of orthostatic hypotension is typically performed through orthostatic testing, which involves repeated measurements of blood pressure and heart rate in supine and standing positions. The tilt-table test may also be employed. However, the advantage of this assessment tool over conventional orthostatic testing is limited. Tilt-table testing is primarily indicated when standing is unsafe, measurements must be standardized, or conditions such as postural orthostatic tachycardia syndrome or neurocardiogenic syncope require evaluation.[55]
Patients with dysautonomia are particularly susceptible to hypotension during anesthesia.[56] This condition may be managed with low doses of phenylephrine, an α1-adrenergic agonist. Supine hypertension in these patients may be treated using transdermal or intravenous nitrates.[57][58]
The SNS contributes to nociception. Evidence suggests that the ANS exerts a regulatory inhibitory effect on pain, and loss of this modulation may create a positive feedback loop, resulting in hyperexcitability of nociceptive fibers. The observation that the analgesic effects of sympathetic nerve blocks often outlast the pharmacologic duration of the anesthetic agents supports this mechanism.
Local sympathetic nerve blocks have been employed to manage select, less common pain conditions, such as complex regional pain syndrome in carefully chosen patients, although evidence for efficacy in phantom limb or postherpetic pain remains limited. Visceral pain may be addressed via a more central approach, including celiac plexus blockade. Given the broad physiological roles of the ANS, sympathetic blocks are generally reserved for intractable pain that is unresponsive to conventional analgesics.[59] For further details, refer to the related StatPearls articles.[60][61][62]
Most conditions related to the ENS are congenital in origin and present during early childhood. The ENS contains excitatory and inhibitory motor neurons. Loss of inhibitory innervation, as observed in life-threatening conditions such as Hirschsprung disease, leads to tonic contraction and obstruction. Presenting complaints often include gastroesophageal reflux, dyspeptic syndromes, constipation, chronic abdominal pain, and irritable bowel syndrome. This condition reflects a failure of embryologic ENS cells to colonize the distal bowel. When the ENS is absent (aganglionosis) or maldeveloped, children experience early constipation, vomiting, progressive growth failure, and, potentially, death.
Studies have identified more than 10 genes with causal associations to Hirschsprung disease, including RET, EDNRB/EDN3, GDNF, SOX10, and PHOX2B. Down syndrome is the most common genetic disorder predisposing individuals to Hirschsprung disease. The chromosome 21-related mechanisms remain incompletely understood.
Media
(Click Image to Enlarge)
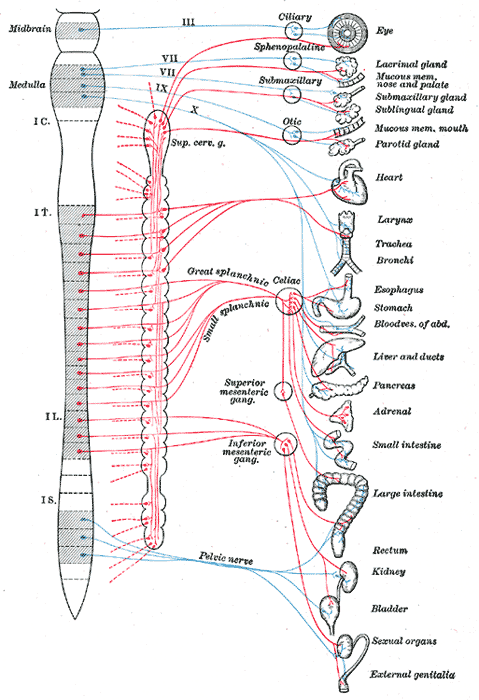
Autonomic Nervous System Pathways. The diagram shows parasympathetic (blue) and sympathetic (red) outflows from the central nervous system to target organs, including ocular, salivary, thoracic, abdominal, and pelvic structures.
Contributed by Henry Gray (1918): Anatomy of the Human Body
(Click Image to Enlarge)
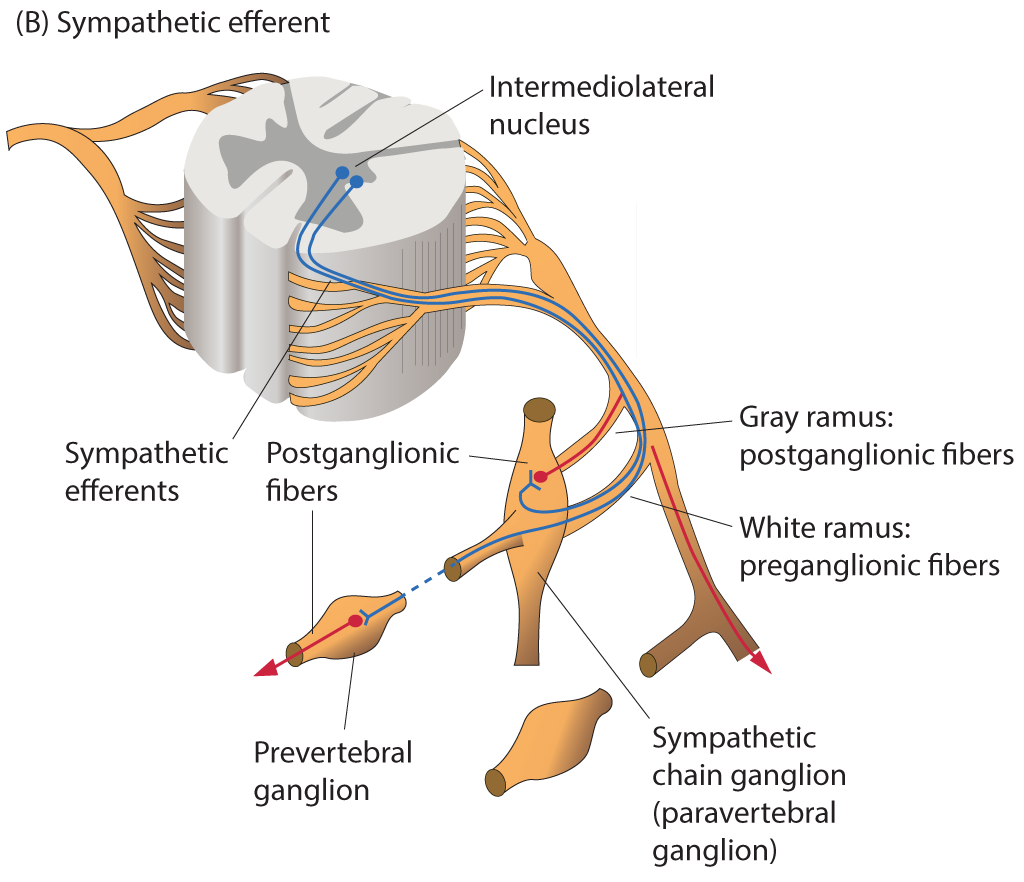
Organization of Sympathetic Efferents. The diagram depicts the intermediolateral nucleus, sympathetic efferents, white and gray rami, sympathetic chain ganglion, postganglionic fibers, and prevertebral ganglion.
Contributed by B Parker
(Click Image to Enlarge)
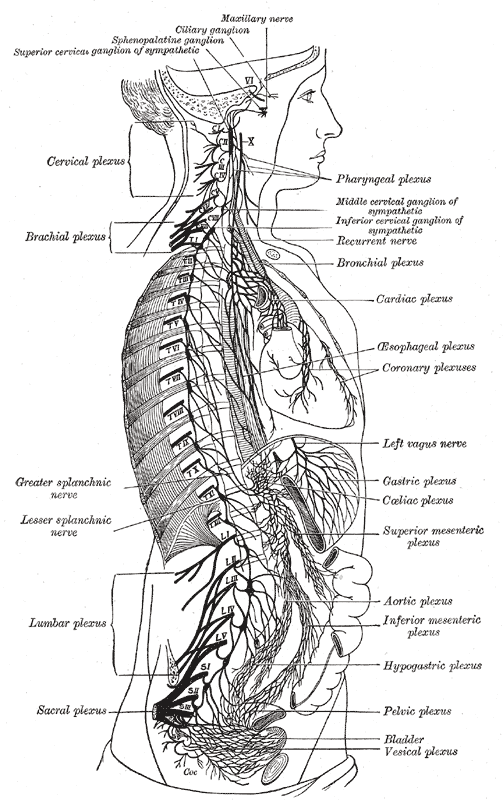
The Sympathetic Nerves. The illustration shows the right sympathetic chain and its connections with the thoracic, abdominal, and pelvic plexuses.
Henry Vandyke Carter, Public Domain, via Wikimedia Commons
(Click Image to Enlarge)
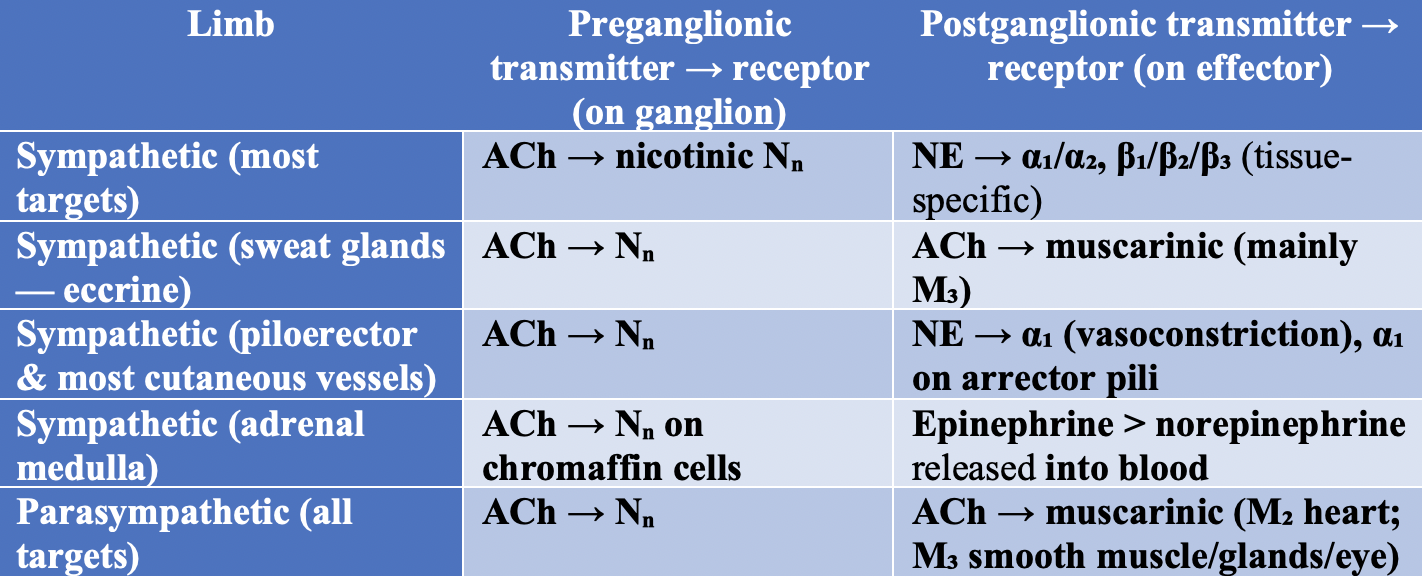
Neurotransmitters of the Autonomic Nervous System. The table summarizes neurotransmitters involved in preganglionic and postganglionic transmission across sympathetic and parasympathetic limbs.
Contributed by Joe M Das, MD. Created using LLM AI.
References
Karemaker JM. An introduction into autonomic nervous function. Physiological measurement. 2017 May:38(5):R89-R118. doi: 10.1088/1361-6579/aa6782. Epub 2017 Mar 17 [PubMed PMID: 28304283]
Siéssere S, Vitti M, Sousa LG, Semprini M, Iyomasa MM, Regalo SC. Anatomic variation of cranial parasympathetic ganglia. Brazilian oral research. 2008 Apr-Jun:22(2):101-5 [PubMed PMID: 18622477]
Koopman FA, Stoof SP, Straub RH, Van Maanen MA, Vervoordeldonk MJ, Tak PP. Restoring the balance of the autonomic nervous system as an innovative approach to the treatment of rheumatoid arthritis. Molecular medicine (Cambridge, Mass.). 2011 Sep-Oct:17(9-10):937-48. doi: 10.2119/molmed.2011.00065. Epub 2011 May 20 [PubMed PMID: 21607292]
Level 3 (low-level) evidenceKenney MJ, Ganta CK. Autonomic nervous system and immune system interactions. Comprehensive Physiology. 2014 Jul:4(3):1177-200. doi: 10.1002/cphy.c130051. Epub [PubMed PMID: 24944034]
Level 3 (low-level) evidenceScott GD, Fryer AD. Role of parasympathetic nerves and muscarinic receptors in allergy and asthma. Chemical immunology and allergy. 2012:98():48-69. doi: 10.1159/000336498. Epub 2012 Jun 26 [PubMed PMID: 22767057]
Level 3 (low-level) evidenceLake JI, Heuckeroth RO. Enteric nervous system development: migration, differentiation, and disease. American journal of physiology. Gastrointestinal and liver physiology. 2013 Jul 1:305(1):G1-24. doi: 10.1152/ajpgi.00452.2012. Epub 2013 May 2 [PubMed PMID: 23639815]
Level 3 (low-level) evidenceSternini C. Organization of the peripheral nervous system: autonomic and sensory ganglia. The journal of investigative dermatology. Symposium proceedings. 1997 Aug:2(1):1-7 [PubMed PMID: 9487007]
Level 3 (low-level) evidenceMcConalogue K, Furness JB. Gastrointestinal neurotransmitters. Bailliere's clinical endocrinology and metabolism. 1994 Jan:8(1):51-76 [PubMed PMID: 7907863]
Level 3 (low-level) evidenceSheehan D, Pick J. The rami communicantes in the rhesus monkey. Journal of anatomy. 1943 Jan:77(Pt 2):125-39 [PubMed PMID: 17104919]
Loukas M, Klaassen Z, Merbs W, Tubbs RS, Gielecki J, Zurada A. A review of the thoracic splanchnic nerves and celiac ganglia. Clinical anatomy (New York, N.Y.). 2010 Jul:23(5):512-22. doi: 10.1002/ca.20964. Epub [PubMed PMID: 20235178]
Yang HJ, Gil YC, Lee WJ, Kim TJ, Lee HY. Anatomy of thoracic splanchnic nerves for surgical resection. Clinical anatomy (New York, N.Y.). 2008 Mar:21(2):171-7. doi: 10.1002/ca.20599. Epub [PubMed PMID: 18288763]
Beveridge TS, Johnson M, Power A, Power NE, Allman BL. Anatomy of the nerves and ganglia of the aortic plexus in males. Journal of anatomy. 2015 Jan:226(1):93-103. doi: 10.1111/joa.12251. Epub 2014 Nov 9 [PubMed PMID: 25382240]
Ehrhardt JD, Weber C, Carey FJ, Lopez-Ojeda W. Anatomy, Thorax, Greater Splanchnic Nerves. StatPearls. 2025 Jan:(): [PubMed PMID: 29763202]
Brindley RL, Bauer MB, Blakely RD, Currie KPM. Serotonin and Serotonin Transporters in the Adrenal Medulla: A Potential Hub for Modulation of the Sympathetic Stress Response. ACS chemical neuroscience. 2017 May 17:8(5):943-954. doi: 10.1021/acschemneuro.7b00026. Epub 2017 Apr 13 [PubMed PMID: 28406285]
Nance DM, Sanders VM. Autonomic innervation and regulation of the immune system (1987-2007). Brain, behavior, and immunity. 2007 Aug:21(6):736-45 [PubMed PMID: 17467231]
Level 3 (low-level) evidenceAlhayek S, Preuss CV. Beta 1 Receptors. StatPearls. 2025 Jan:(): [PubMed PMID: 30422499]
Farzam K, Kidron A, Lakhkar AD. Adrenergic Drugs. StatPearls. 2025 Jan:(): [PubMed PMID: 30480963]
Khalid MM, Galuska MA, Hamilton RJ. Beta-Blocker Toxicity. StatPearls. 2025 Jan:(): [PubMed PMID: 28846217]
Heusch G, Thämer V. [Significance of the sympathetic nervous system for the coronary circulation]. Zeitschrift fur Kardiologie. 1984 Sep:73(9):543-51 [PubMed PMID: 6506839]
Vargas Pelaez AF, Gao Z, Ahmad TA, Leuenberger UA, Proctor DN, Maman SR, Muller MD. Effect of adrenergic agonists on coronary blood flow: a laboratory study in healthy volunteers. Physiological reports. 2016 May:4(10):. doi: 10.14814/phy2.12806. Epub [PubMed PMID: 27225628]
Chen IC, Kuo J, Ko WJ, Shih HC, Kuo CD. Increased flow resistance and decreased flow rate in patients with acute respiratory distress syndrome: The role of autonomic nervous modulation. Journal of the Chinese Medical Association : JCMA. 2016 Jan:79(1):17-24. doi: 10.1016/j.jcma.2015.10.001. Epub 2015 Nov 14 [PubMed PMID: 26589196]
Sternberg EM. Neural regulation of innate immunity: a coordinated nonspecific host response to pathogens. Nature reviews. Immunology. 2006 Apr:6(4):318-28 [PubMed PMID: 16557263]
Level 3 (low-level) evidenceElenkov IJ, Wilder RL, Chrousos GP, Vizi ES. The sympathetic nerve--an integrative interface between two supersystems: the brain and the immune system. Pharmacological reviews. 2000 Dec:52(4):595-638 [PubMed PMID: 11121511]
Level 3 (low-level) evidenceBellinger DL, Millar BA, Perez S, Carter J, Wood C, ThyagaRajan S, Molinaro C, Lubahn C, Lorton D. Sympathetic modulation of immunity: relevance to disease. Cellular immunology. 2008 Mar-Apr:252(1-2):27-56. doi: 10.1016/j.cellimm.2007.09.005. Epub 2008 Mar 4 [PubMed PMID: 18308299]
Level 3 (low-level) evidenceLundberg JM, Hökfelt T. Multiple co-existence of peptides and classical transmitters in peripheral autonomic and sensory neurons--functional and pharmacological implications. Progress in brain research. 1986:68():241-62 [PubMed PMID: 2882554]
Level 3 (low-level) evidenceLundberg JM, Hökfelt T, Schultzberg M, Uvnäs-Wallensten K, Köhler C, Said SI. Occurrence of vasoactive intestinal polypeptide (VIP)-like immunoreactivity in certain cholinergic neurons of the cat: evidence from combined immunohistochemistry and acetylcholinesterase staining. Neuroscience. 1979:4(11):1539-59 [PubMed PMID: 390416]
Level 3 (low-level) evidenceLandis SC, Fredieu JR. Coexistence of calcitonin gene-related peptide and vasoactive intestinal peptide in cholinergic sympathetic innervation of rat sweat glands. Brain research. 1986 Jul 2:377(1):177-81 [PubMed PMID: 3524749]
Level 3 (low-level) evidenceLindh B, Lundberg JM, Hökfelt T, Elfvin LG, Fahrenkrug J, Fischer J. Coexistence of CGRP- and VIP-like immunoreactivities in a population of neurons in the cat stellate ganglia. Acta physiologica Scandinavica. 1987 Nov:131(3):475-6 [PubMed PMID: 3321916]
Level 3 (low-level) evidenceBrain SD, Williams TJ, Tippins JR, Morris HR, MacIntyre I. Calcitonin gene-related peptide is a potent vasodilator. Nature. 1985 Jan 3-9:313(5997):54-6 [PubMed PMID: 3917554]
Level 3 (low-level) evidenceBerthoud HR, Neuhuber WL. Functional and chemical anatomy of the afferent vagal system. Autonomic neuroscience : basic & clinical. 2000 Dec 20:85(1-3):1-17 [PubMed PMID: 11189015]
Level 3 (low-level) evidencePavlov VA, Wang H, Czura CJ, Friedman SG, Tracey KJ. The cholinergic anti-inflammatory pathway: a missing link in neuroimmunomodulation. Molecular medicine (Cambridge, Mass.). 2003 May-Aug:9(5-8):125-34 [PubMed PMID: 14571320]
Level 3 (low-level) evidenceTsuchihashi K, Yoshihiro T, Aikawa T, Nio K, Takayoshi K, Yokoyama T, Fukata M, Arita S, Ariyama H, Shimizu Y, Yoshida Y, Torisu T, Esaki M, Odashiro K, Kusaba H, Akashi K, Baba E. Metastatic esophageal cancer presenting as shock by injury of vagus nerve mimicking baroreceptor reflex: A case report. Medicine. 2017 Dec:96(49):e8987. doi: 10.1097/MD.0000000000008987. Epub [PubMed PMID: 29245271]
Level 3 (low-level) evidenceWood JD. Application of classification schemes to the enteric nervous system. Journal of the autonomic nervous system. 1994 Jun:48(1):17-29 [PubMed PMID: 8027516]
Level 3 (low-level) evidenceThayer JF, Sternberg EM. Neural aspects of immunomodulation: focus on the vagus nerve. Brain, behavior, and immunity. 2010 Nov:24(8):1223-8. doi: 10.1016/j.bbi.2010.07.247. Epub 2010 Jul 30 [PubMed PMID: 20674737]
Level 3 (low-level) evidenceLundberg JM, Hökfelt T, Anggård A, Uvnäs-Wallensten K, Brimijoin S, Brodin E, Fahrenkrug J. Peripheral peptide neurons: distribution, axonal transport, and some aspects on possible function. Advances in biochemical psychopharmacology. 1980:22():25-36 [PubMed PMID: 6156578]
Level 3 (low-level) evidenceLeblanc GG, Trimmer BA, Landis SC. Neuropeptide Y-like immunoreactivity in rat cranial parasympathetic neurons: coexistence with vasoactive intestinal peptide and choline acetyltransferase. Proceedings of the National Academy of Sciences of the United States of America. 1987 May:84(10):3511-5 [PubMed PMID: 3554241]
Level 3 (low-level) evidenceCarlson AB, Kraus GP. Physiology, Cholinergic Receptors. StatPearls. 2025 Jan:(): [PubMed PMID: 30252390]
Million M, Larauche M. Stress, sex, and the enteric nervous system. Neurogastroenterology and motility. 2016 Sep:28(9):1283-9. doi: 10.1111/nmo.12937. Epub [PubMed PMID: 27561694]
Wood JD. Enteric Nervous System: Neuropathic Gastrointestinal Motility. Digestive diseases and sciences. 2016 Jul:61(7):1803-16. doi: 10.1007/s10620-016-4183-5. Epub [PubMed PMID: 27142673]
Hansen MB. The enteric nervous system I: organisation and classification. Pharmacology & toxicology. 2003 Mar:92(3):105-13 [PubMed PMID: 12753424]
Level 3 (low-level) evidenceSarna SK. Colonic Motility: From Bench Side to Bedside. 2010:(): [PubMed PMID: 21452445]
Butler SJ, Bronner ME. From classical to current: analyzing peripheral nervous system and spinal cord lineage and fate. Developmental biology. 2015 Feb 15:398(2):135-46. doi: 10.1016/j.ydbio.2014.09.033. Epub 2014 Oct 24 [PubMed PMID: 25446276]
Level 3 (low-level) evidenceDyachuk V, Furlan A, Shahidi MK, Giovenco M, Kaukua N, Konstantinidou C, Pachnis V, Memic F, Marklund U, Müller T, Birchmeier C, Fried K, Ernfors P, Adameyko I. Neurodevelopment. Parasympathetic neurons originate from nerve-associated peripheral glial progenitors. Science (New York, N.Y.). 2014 Jul 4:345(6192):82-7. doi: 10.1126/science.1253281. Epub 2014 Jun 12 [PubMed PMID: 24925909]
Level 3 (low-level) evidenceGariepy CE. Intestinal motility disorders and development of the enteric nervous system. Pediatric research. 2001 May:49(5):605-13 [PubMed PMID: 11328941]
Level 3 (low-level) evidenceSasselli V, Pachnis V, Burns AJ. The enteric nervous system. Developmental biology. 2012 Jun 1:366(1):64-73. doi: 10.1016/j.ydbio.2012.01.012. Epub 2012 Jan 24 [PubMed PMID: 22290331]
Level 3 (low-level) evidenceDiamantis E, Farmaki P, Savvanis S, Athanasiadis G, Troupis T, Damaskos C. Sympathetic Nerve Injury in Thyroid Cancer. Acta medica (Hradec Kralove). 2017:60(4):135-139. doi: 10.14712/18059694.2018.8. Epub [PubMed PMID: 29716678]
Kanagalingam S, Miller NR. Horner syndrome: clinical perspectives. Eye and brain. 2015:7():35-46. doi: 10.2147/EB.S63633. Epub 2015 Apr 10 [PubMed PMID: 28539793]
Level 3 (low-level) evidenceKhan Z, Bollu PC. Horner Syndrome. StatPearls. 2025 Jan:(): [PubMed PMID: 29763176]
Lykstad J, Reddy V, Hanna A. Neuroanatomy, Pupillary Dilation Pathway. StatPearls. 2026 Jan:(): [PubMed PMID: 30571042]
Delort S, Marchi E, Corrêa MA. Oxybutynin as an alternative treatment for hyperhidrosis. Anais brasileiros de dermatologia. 2017 Mar-Apr:92(2):217-220. doi: 10.1590/abd1806-4841.201755126. Epub [PubMed PMID: 28538882]
del Boz J. Systemic treatment of hyperhidrosis. Actas dermo-sifiliograficas. 2015 May:106(4):271-7. doi: 10.1016/j.ad.2014.11.012. Epub 2015 Jan 28 [PubMed PMID: 25638324]
Haam SJ, Park SY, Paik HC, Lee DY. Sympathetic nerve reconstruction for compensatory hyperhidrosis after sympathetic surgery for primary hyperhidrosis. Journal of Korean medical science. 2010 Apr:25(4):597-601. doi: 10.3346/jkms.2010.25.4.597. Epub 2010 Mar 19 [PubMed PMID: 20358004]
McLeod JG. Investigation of peripheral neuropathy. Journal of neurology, neurosurgery, and psychiatry. 1995 Mar:58(3):274-83 [PubMed PMID: 7897405]
Ludwig PE, Reddy V, Varacallo MA. Neuroanatomy, Central Nervous System (CNS)(Archived). StatPearls. 2025 Jan:(): [PubMed PMID: 28723039]
Mustafa HI, Fessel JP, Barwise J, Shannon JR, Raj SR, Diedrich A, Biaggioni I, Robertson D. Dysautonomia: perioperative implications. Anesthesiology. 2012 Jan:116(1):205-15. doi: 10.1097/ALN.0b013e31823db712. Epub [PubMed PMID: 22143168]
Sánchez-Manso JC, Gujarathi R, Varacallo MA. Autonomic Dysfunction. StatPearls. 2025 Jan:(): [PubMed PMID: 28613638]
Bevan DR. Shy-Drager syndrome. A review and a description of the anaesthetic management. Anaesthesia. 1979 Oct:34(9):866-73 [PubMed PMID: 532923]
Level 3 (low-level) evidenceStirt JA, Frantz RA, Gunz EF, Conolly ME. Anesthesia, catecholamines, and hemodynamics in autonomic dysfunction. Anesthesia and analgesia. 1982 Aug:61(8):701-4 [PubMed PMID: 7201274]
Level 3 (low-level) evidenceGunduz OH, Kenis-Coskun O. Ganglion blocks as a treatment of pain: current perspectives. Journal of pain research. 2017:10():2815-2826. doi: 10.2147/JPR.S134775. Epub 2017 Dec 14 [PubMed PMID: 29276402]
Level 3 (low-level) evidenceJOHN RS, Dixon B, Hendrix JM, Shienbaum R. Celiac Plexus Block. StatPearls. 2025 Jan:(): [PubMed PMID: 30285364]
Piraccini E, Munakomi S, Chang KV. Stellate Ganglion Blocks. StatPearls. 2025 Jan:(): [PubMed PMID: 29939575]
Alexander CE, De Jesus O, Varacallo MA. Lumbar Sympathetic Block. StatPearls. 2025 Jan:(): [PubMed PMID: 28613759]