Introduction
Genetic mosaicism is characterized by the presence of 2 or more genetically distinct cell populations within an individual, all originating from a single zygote. Mosaicism results from postzygotic genetic alterations that occur after fertilization. In contrast, chimerism arises from the coexistence of cell populations derived from 2 or more distinct zygotes.
The human body undergoes an estimated 1016 cell divisions during a lifetime. Because each mitotic event carries a risk of genetic error, virtually all individuals harbor genetically distinct cell populations and are therefore mosaic to some degree. Although most postzygotic mutations are clinically silent or are eliminated by mechanisms such as apoptosis, some persist and contribute to disease. Somatic mosaicism has been implicated in a wide range of human pathologies, including chromosomal disorders such as Turner Syndrome and the development of benign and malignant neoplasms, which are themselves manifestations of mosaicism.[1][2][3][4][5]
Development
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Development
Human development begins with the formation of a zygote through the fusion of a haploid sperm and ovum, each containing 23 chromosomes. As the zygote undergoes repeated mitotic divisions to generate the cells of the body, postzygotic genetic alterations may occur. Consequently, mosaicism can develop at any stage after zygote formation, resulting in 2 or more genetically distinct cell populations within a single individual.[1][2]
Mosaicism is broadly classified into 2 categories:
- Somatic mosaicism: This form occurs when 2 or more genetically distinct cell lineages are present within somatic tissues. Because germ cells (sperm and oocytes) are generally unaffected, somatic mosaicism is not typically transmitted to offspring. However, recent evidence suggests that somatic mosaicism arising during the preimplantation stage of embryonic development may involve both somatic and germline cell lineages, creating the potential for transmission to future generations.[3][4]
- Germline mosaicism: This form occurs when 2 or more genetically distinct cell lineages are present within germ cells. Unlike somatic mosaicism, germline mosaicism can be transmitted to offspring. Individuals with germline mosaicism may be clinically unaffected if the pathogenic variant is confined to germ cells; however, the mutation may be inherited by offspring, who may then manifest the associated genetic disorder.[5][6][7]
Constitutional mosaicism occurs during the preimplantation or embryonic stages of development and becomes an integral part of the organism. In contrast, somatic mosaicism results from postzygotic genetic alterations that occur after embryogenesis and is generally restricted to specific tissues or cell populations.[8][9] Constitutional mosaicism results from postzygotic errors in chromosome segregation during mitotic cell division early in embryonic development.[10]
Mosaic embryos may be classified into 3 categories (Image 1):
- Aneuploid mosaic: Aneuploid mosaicism consists of multiple aneuploid cell lines within the same embryo, with all affected cell populations exhibiting abnormal chromosome numbers.[11] IIn chromosomal aneuploid mosaicism, cells may lack an entire chromosome (monosomy; 45 chromosomes) or contain an extra chromosome (trisomy; 47 chromosomes).[12][13]
- Euploid-aneuploid mosaic: Euploid-aneuploid mosaicism is characterized by the coexistence of euploid and aneuploid cell populations within the same embryo. Some cells contain the normal complement of 46 chromosomes, whereas others are aneuploid (eg, 45 or 47 chromosomes). Clinical severity is influenced by the proportion and distribution of euploid and aneuploid cells. This form of mosaicism is relatively common and is associated with a greater likelihood of embryonic survival.[6][14]
- Ploidy mosaics: Ploidy mosaicism, also known as mixoploidy, involves the presence of cell populations with different multiples of the haploid chromosome number.[15] In these cases, an additional complete set of chromosomes is present in affected cells, as seen in triploidy (69 chromosomes) or tetraploidy (92 chromosomes). Embryos with predominantly triploid or tetraploid cell populations are generally nonviable. However, triploid and tetraploid cells may occur physiologically in certain tissues, including the liver and bone marrow.[16][17][15]
The distribution of mosaicism and its phenotypic manifestations depend largely on the timing of the mutation during embryonic development. If a mutation occurs during the first mitotic division after fertilization, approximately half of the individual's cells will carry the mutation, potentially producing a distinct midline demarcation between affected and unaffected tissues. Although the precise timing of left-right axis separation in humans remains uncertain, mutations arising after this event are more likely to be confined to one side of the body and less likely to cross the midline.
Mutations that occur before primordial germ cell differentiation (approximately within the first 15 mitotic divisions) may be present in both somatic and germline tissues. In contrast, mutations arising after this stage are generally restricted to either somatic or germline cell lineages. Consequently, estimating the timing of a mutation can help predict the proportion and distribution of mutant cells and may provide insight into the risk of transmitting the mutation to future offspring.[18]
Cellular
Based on the distribution of affected cells, mosaicism can be classified into 2 broad categories:
- Generalized mosaicism: In generalized mosaicism, 2 or more genetically distinct cell populations are distributed throughout the organism. This form of mosaicism typically results from a mutation occurring before cellular differentiation, allowing the abnormal cell line to contribute to multiple tissues and organ systems. Generalized mosaicism may also arise from mutations present in paternal or maternal germ cells that are transmitted to the zygote.[13][19][20]
- Confined mosaicism: In confined mosaicism, genetically distinct cell populations are restricted to specific tissues, organs, or regions of the body rather than being distributed throughout the organism. Examples include mosaicism limited to the brain, heart, or liver. A specialized form, confined placental mosaicism, occurs when chromosomal abnormalities are present in placental tissues but absent from the fetus and may arise during early embryonic development or placental growth.[21][22]
The consequences of mosaicism depend on the specific genetic alteration involved. Genetically distinct cell populations may be phenotypically indistinguishable when the variant does not alter gene expression or cellular function. Such cases may occur when the mutation affects a noncoding genomic region or when a recessive variant is not expressed in the presence of a normal allele.[23][24]
Mosaicism arises from postzygotic genetic alterations, including single-nucleotide variants, chromosomal abnormalities, and copy number variants, that result in genetically distinct cell populations within an individual. The extent and clinical impact of mosaicism depend on the developmental timing of the mutation and the proportion and distribution of affected cells. Mosaicism may be limited to somatic tissues, confined to the germline, or involve both somatic and germline lineages. In some cases, germline mosaicism can result in transmission of the mutation to offspring despite the absence of clinical manifestations in the parent.[4][25]
The phenotypic expression of mosaicism depends on the proportion and distribution of affected cells. Clinical manifestations may arise during intrauterine development, at birth, or later in life, depending on the specific genetic alteration and other modifying factors.[26][27]
Biochemical
Mosaicism may result from gene-level mutations arising through the following mechanisms:
- Single-nucleotide variants
- Small insertions or deletions
- Trinucleotide repeat expansions and contractions
- Autonomous mobile element insertions [18]
Chromosomal and gene-level mutations may alter the biochemical activity of affected cells by changing the quantity, structure, or function of expressed gene products. For example, expression of myelin basic protein in the optic nerve and spinal cord is altered in heterozygous female mice carrying the rumpshaker mutation, an X-linked genetic defect.[28] Similarly, chromosomal abnormalities such as tetrasomy 3q26.32-q29 have been associated with cutaneous hyperpigmentation.[29] In X-linked Agammaglobulinemia, affected individuals fail to develop mature B lymphocytes, resulting in an absence or severe deficiency of plasma cells and markedly reduced antibody production.[30]
Some studies have reported an association between mosaic Trisomy 13 and hypomelanosis of Ito, a condition characterized by localized areas of hypopigmentation resulting from abnormal melanocyte function or distribution.[31]
Molecular Level
The early embryo is particularly vulnerable to mitotic errors because embryonic genome activation occurs only after the first several cell divisions. During this period, development depends primarily on maternally derived transcripts and proteins within the oocyte. Delayed expression of genes involved in cell division and chromosome segregation contributes to the high frequency of chromosomal mosaicism observed during early embryogenesis. Accordingly, mosaicism has been reported in up to 70% of cleavage-stage embryos and 90% of blastocyst-stage embryos generated through in vitro fertilization.[32][13]
Function
The functional consequences of mosaicism depend largely on the proportion, distribution, and type of affected cells. Clinical manifestations may vary considerably, as illustrated by the following examples:
- Mosaic loss of the Y chromosome in hematopoietic cells has been associated with increased morbidity and mortality in older adults.[33]
- Mosaic Klinefelter Syndrome (46,XY/47,XXY) is associated with reduced testicular volume and impaired testosterone production.[34]
- In sporadic retinoblastoma, both the degree and distribution of mosaicism influence disease severity and age at onset.[35]
- Mosaic variants in the SMC1A gene have been identified in buccal mucosal cells of individuals with clinically diagnosed Cornelia de Lange Syndrome.[36]
In some cases, mosaicism has little or no functional consequence. This may occur when the mutation is phenotypically silent, affects a recessive allele, or is present in too few cells to produce a detectable clinical effect. A classic example involves X-linked disorders.
In X-linked conditions, the clinical phenotype may be modified by mosaicism. For example, pathogenic variants in the MECP2 gene, which cause Rett Syndrome, are typically lethal in hemizygous males because they possess only a single X chromosome. However, if a male is mosaic for a pathogenic MECP2 variant, a proportion of cells retain a normal copy of the gene. The presence of these unaffected cells may mitigate disease severity and increase the likelihood of survival.[37]
Some evidence suggests that mosaicism may serve as a protective mechanism, allowing survival in the presence of chromosomal abnormalities that would otherwise be incompatible with life. Only a limited number of full autosomal trisomies, most notably trisomies 13, 18, and 21, are compatible with live birth. In contrast, a broader spectrum of chromosomal abnormalities has been documented in the mosaic state. Reported examples include mosaic trisomies involving chromosomes 8 (Warkany syndrome), 14, 16, and 17. Likewise, complete autosomal monosomies are generally considered incompatible with human survival, with only rare reports of mosaic monosomy 21. Another notable example is mosaic tetrasomy 12p, which causes Pallister-Killian Syndrome. In such conditions, survival is thought to be possible because a sufficient proportion of cells retain a normal chromosomal complement, thereby mitigating the effects of the chromosomal abnormality.[38][18][39]
Mechanism
Somatic mosaicism may arise from a wide range of genetic alterations, including chromosomal abnormalities, copy number variants, and single-nucleotide mutations. Mosaic aneuploidies are generated primarily through 2 mechanisms (see Image 2): postzygotic mitotic nondisjunction and postzygotic trisomy rescue following meiotic nondisjunction. The latter mechanism represents an attempt to restore a normal chromosomal complement through the loss of a supernumerary chromosome, a process known as chromosomal rescue. Although chromosomal rescue may reestablish euploidy, it can also result in uniparental disomy, which may disrupt genomic imprinting and increase the risk of homozygosity for pathogenic recessive variants.[38][18][40] Additional mechanisms that can give rise to mosaicism include anaphase lag and endoreduplication.[13]
Testing
The detection of embryonic and fetal mosaicism has become increasingly feasible through prenatal diagnostic testing. The most commonly used invasive prenatal diagnostic procedures are chorionic villus sampling and amniocentesis, each with distinct advantages and limitations.[8]
- Amniocentesis: Amniotic fluid is aspirated from the uterine cavity, typically at or after 15 weeks of gestation, for cytogenetic and molecular genetic analysis. Amniocentesis performed before 15 weeks of gestation is associated with an increased risk of fetal complications and is generally not recommended.[41][42]
- Chorionic villus sampling: Chorionic villus tissue is obtained for genetic analysis, typically beginning at approximately 10 weeks of gestation. Because the sampled tissue is of placental origin, results may occasionally reflect confined placental mosaicism rather than the fetal genotype.[41]
Neither procedure directly samples embryonic or fetal tissue; instead, genetic analysis is performed on amniotic fluid cells or placental tissue obtained through chorionic villus sampling.
Multiple laboratory techniques have been developed to detect mosaicism and other chromosomal abnormalities.
Karyotyping
Karyotyping is a conventional cytogenetic technique that can identify different chromosomal complements within a single individual. It remains widely used in clinical practice and enables detection of numerical and large structural chromosomal abnormalities, including aneuploidies, translocations, and large deletions or duplications, typically greater than 5 Mb. However, its resolution is limited, and low-level mosaicism may be difficult to detect.[43]
Fluorescent In Situ Hybridization
Fluorescence in situ hybridization (FISH) enables targeted analysis of specific chromosomal regions and can detect smaller copy number changes than conventional karyotyping, often at resolutions approaching 50 kb. Because FISH can be performed on interphase nuclei, it allows evaluation of large numbers of cells and may improve the detection of mosaicism. Nevertheless, accurate identification of low-level mosaicism often requires analysis of a substantial number of cells.[44][45]
Sanger Sequencing
Sanger sequencing permits nucleotide-level analysis and remains useful for targeted evaluation of specific genes and variants. However, because it is limited to relatively small genomic regions, it has largely been supplanted by high-throughput genomic technologies that can simultaneously assess large portions of the genome.[38][18][46]
Chromosomal Microarray
Chromosomal microarray (CMA) detects copy number variations across the genome without requiring cell culture. CMA includes both comparative genomic hybridization (CGH) arrays and single-nucleotide polymorphism (SNP) arrays. Compared with conventional cytogenetic techniques, CMA offers improved sensitivity for mosaicism, with reported detection thresholds of approximately 10% to 20% for CGH arrays and as low as 5% for SNP arrays.[43][47][48]
Next-Generation Sequencing
Next-generation sequencing (NGS) provides high-resolution, genome-wide analysis and can identify single-nucleotide variants, small insertions and deletions, copy-number alterations, and other genomic abnormalities. Its high sequencing depth enables detection of low-level mosaicism that may be missed by other methods. NGS is particularly useful for identifying nucleotide-level variants and has become an important tool in clinical genetics, assisted reproductive technology, and preimplantation genetic testing.[38][49]
Pathophysiology
Gene-level and chromosomal alterations may result in genomic instability, producing genetically distinct cell populations within the same individual. Mosaic chromosomal abnormalities can lead to altered cellular function through changes in gene dosage and gene expression, resulting in abnormal protein production and diverse clinical phenotypes.[50]
The clinical consequences of mosaicism are illustrated by several examples:
- Somatic chromosomal abnormalities may influence expression of the amyloid precursor protein and have been implicated in neurodegenerative disorders such as Alzheimer Disease.[51]
- In Down Syndrome, individuals with mosaic trisomy 21 generally exhibit milder cognitive impairment than those with nonmosaic trisomy 21, reflecting the presence of both trisomic (47,XX,+21 or 47,XY,+21) and euploid (46,XX or 46,XY) cell populations.[52]
- In mosaic Turner Syndrome, loss of an X chromosome during early embryonic cell division results in both 45,X and 46,XX cell lines. Individuals with mosaic Turner syndrome often have a milder phenotype than those with complete monosomy X.[53]
The biological consequences of somatic mosaicism can be conceptualized as a sequence of events:
- A postzygotic genetic alteration occurs during mitosis.
- Two or more genetically distinct cell populations develop.
- Cellular function may be altered through changes in gene expression, protein production, or hormone synthesis.
- If a sufficient proportion of cells is affected, tissue and organ function may become impaired.
- Clinical manifestations emerge, with severity depending on the distribution and proportion of affected cells.
- In some cases, expansion of the mutant cell population may contribute to neoplastic transformation and malignancy.[54][55][56][57][58][59][6]
The consequences of germline mosaicism differ because the mutation is confined to germ cells:
- A genetic alteration arises in a subset of germ cells during gametogenesis.
- The affected individual is often clinically unaffected because somatic tissues do not carry the mutation.
- An affected germ cell participates in fertilization, transmitting the variant to the offspring.
- The offspring may inherit the mutation in all cells or as a mosaic state, depending on the timing and mechanism of the genetic alteration.
- Clinical manifestations in the offspring depend on the specific mutation and the proportion and distribution of affected cells.[60][61][62][6][63]
Clinical Significance
Mosaicism has important implications for genetic counseling, particularly when identified during prenatal testing.[64] Because chromosomal nondisjunction occurs more frequently during oogenesis than spermatogenesis, counseling should consider the origin and type of chromosomal abnormality. Although the recurrence risk associated with mosaicism is often difficult to predict, germline (gonadal) mosaicism should be considered when a child is affected by a seemingly de novo genetic disorder or chromosomal abnormality. Germline mosaicism may explain familial recurrence of rare pathogenic variants despite unaffected parents.[65] Because the clinical consequences vary according to the specific chromosome or gene involved, genetic counseling should be individualized based on the nature and extent of the mosaicism.[8]
Mosaicism can influence survival in chromosomal disorders that are typically incompatible with life. Although complete autosomal monosomies are generally lethal in humans, mosaic forms may be compatible with survival when a sufficient proportion of cells retain a normal chromosomal complement.[66][67]
Mosaicism has also been implicated in oncogenesis. Age-related accumulation of somatic genetic alterations contributes to tissue mosaicism and may increase the risk of malignant transformation in susceptible cell populations.[56][57]
Several single-gene disorders demonstrate clinically significant mosaicism:
- Hereditary tyrosinemia type 1: Pathogenic variants in the FAH gene cause hereditary tyrosinemia type 1. Somatic mosaicism may occur in hepatocytes, resulting in populations of both affected and genetically corrected cells.[68][64]
- Bloom syndrome: Pathogenic variants in the BLM gene, which encodes a DNA helicase involved in DNA replication and repair, cause Bloom syndrome, a disorder characterized by growth impairment, immunodeficiency, genomic instability, and increased cancer risk.[69][70]
- Duchenne muscular dystrophy: Mosaicism involving pathogenic variants in the dystrophin gene has been reported in affected individuals and carrier mothers.[71]
Mosaicism is also an important consideration in assisted reproductive technologies. Embryonic mosaicism is relatively common in embryos generated through in vitro fertilization, and many fertility centers incorporate preimplantation genetic testing to identify chromosomal abnormalities and reduce the risk of transmitting genetic disorders.[6][49][19]
Media
(Click Image to Enlarge)
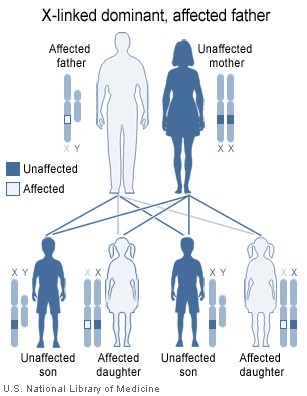
Image showing inheritance of X-linked dominant mutation from an affected father, The sons of a man with an X-linked dominant disorder will not be affected, but his daughters will all inherit the condition Contributed by National Institute of Health ( http://ghr.nlm.nih.gov/handbook/illustrations/xlinkdominantfather )
(Click Image to Enlarge)
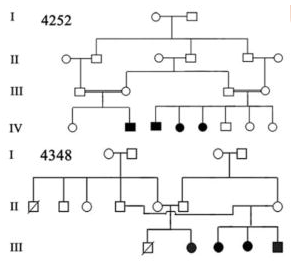
Image 1: Pedigree of a Prevalent Autosomal Recessive Disease Contributed by Chishti, Muhammad S et al. “Splice-site mutations in the TRIC gene underlie autosomal recessive nonsyndromic hearing impairment in Pakistani families.” Journal of human genetics vol. 53,2 (2008): 101-5. doi:10.1007/s10038-007-0209-3 (https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2757049/)
(Click Image to Enlarge)
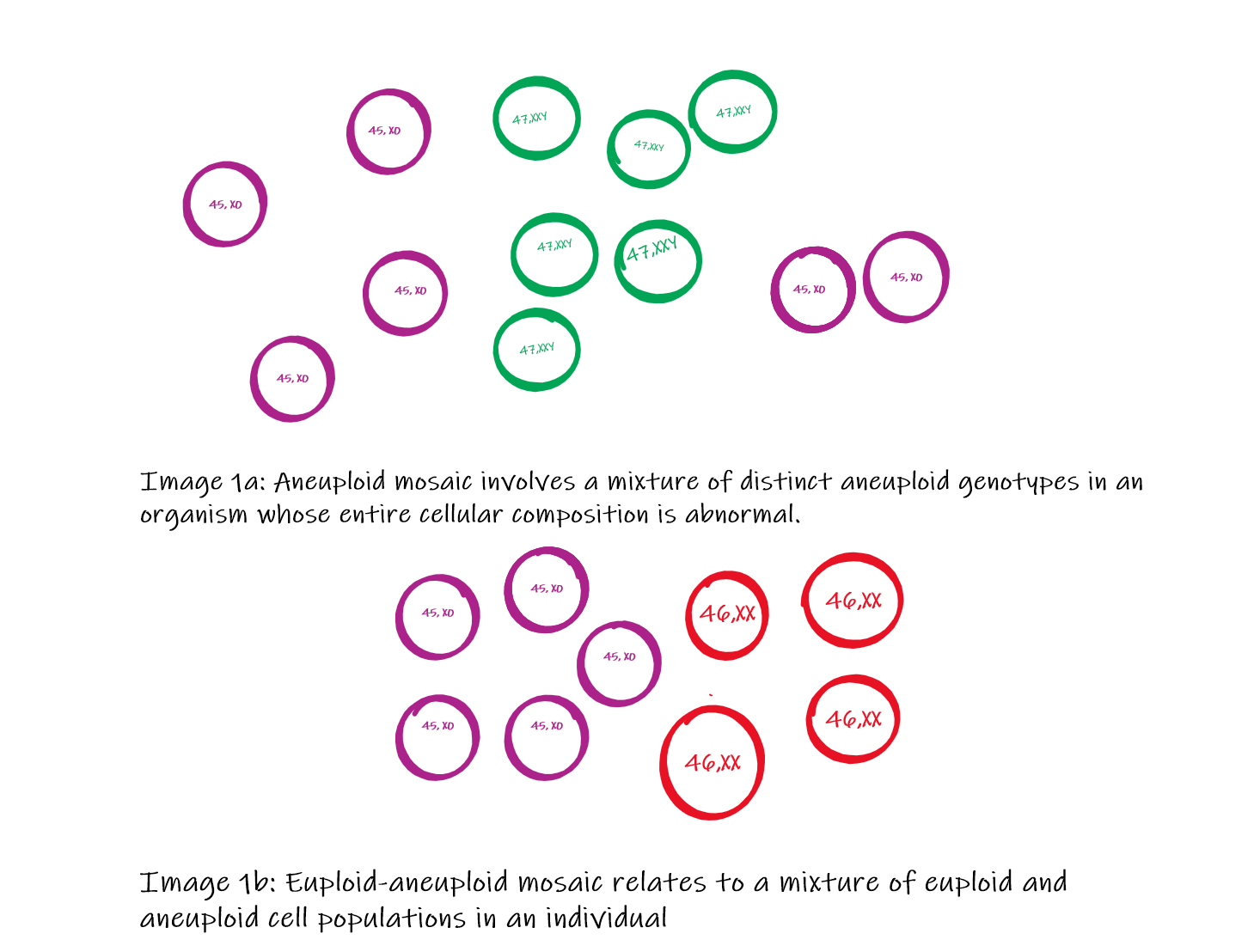
Types of Mosaicism Contributed by Dr. Pradip Chauhan. MS, P.D.U. Government Medical College, Rajkot, Gujarat, India
(Click Image to Enlarge)
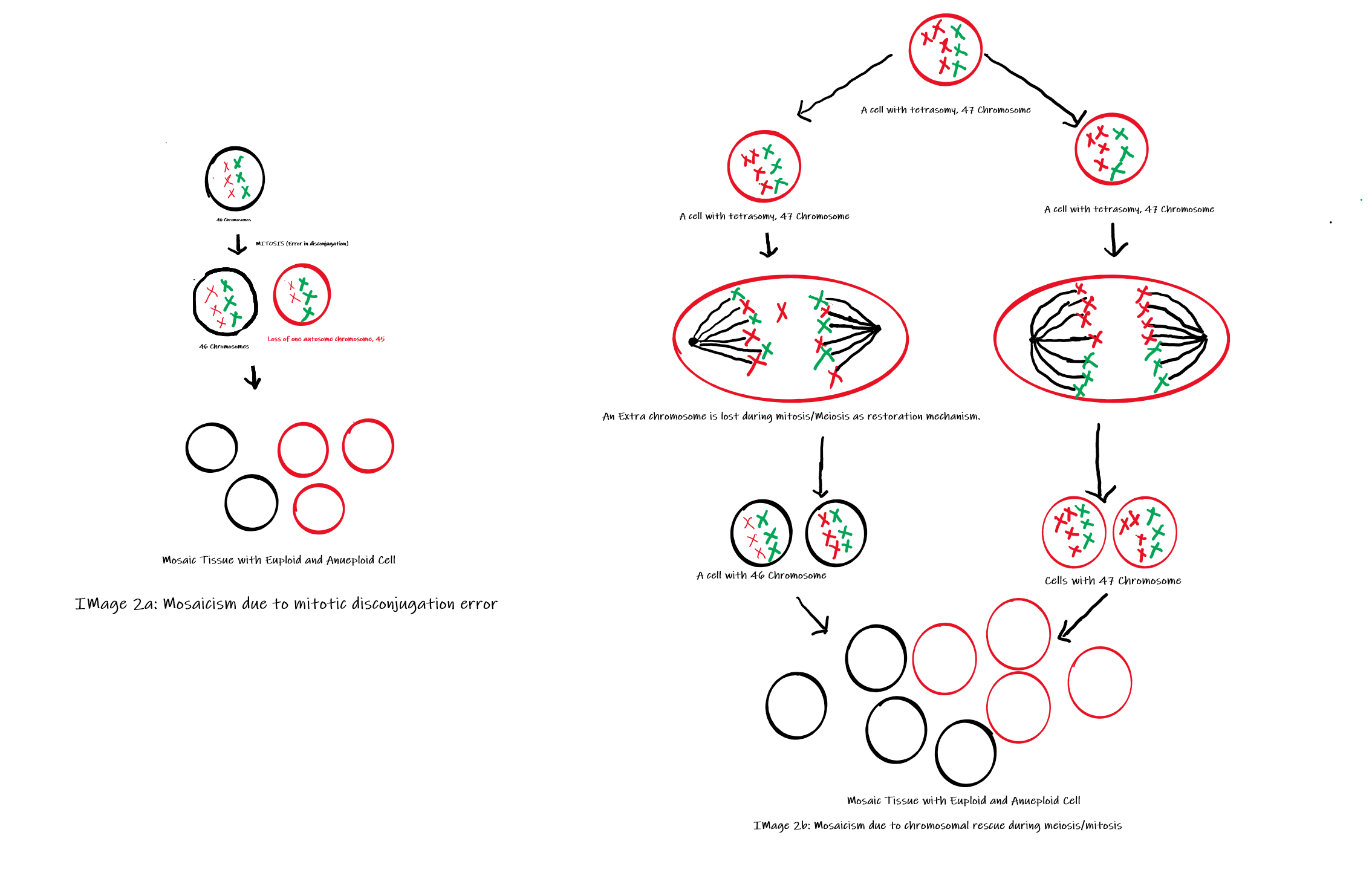
Mechanism of Mosaicism (Image 2a: Shows mosaicism due to disconjugation, Image 2b: Shows mosaicism due to chromosomal rescue mechanism) Contributed by Dr. Pradip Chauhan, MS (Anatomy); P.D.U. Government Medical College, Rajkot, India.
References
Popovic M, Dhaenens L, Boel A, Menten B, Heindryckx B. Chromosomal mosaicism in human blastocysts: the ultimate diagnostic dilemma. Human reproduction update. 2020 Apr 15:26(3):313-334. doi: 10.1093/humupd/dmz050. Epub [PubMed PMID: 32141501]
Foulkes WD, Real FX. Many mosaic mutations. Current oncology (Toronto, Ont.). 2013 Apr:20(2):85-7. doi: 10.3747/co.20.1449. Epub [PubMed PMID: 23559869]
Holstege H, Pfeiffer W, Sie D, Hulsman M, Nicholas TJ, Lee CC, Ross T, Lin J, Miller MA, Ylstra B, Meijers-Heijboer H, Brugman MH, Staal FJ, Holstege G, Reinders MJ, Harkins TT, Levy S, Sistermans EA. Somatic mutations found in the healthy blood compartment of a 115-yr-old woman demonstrate oligoclonal hematopoiesis. Genome research. 2014 May:24(5):733-42. doi: 10.1101/gr.162131.113. Epub 2014 Apr 23 [PubMed PMID: 24760347]
Gajecka M. Unrevealed mosaicism in the next-generation sequencing era. Molecular genetics and genomics : MGG. 2016 Apr:291(2):513-30. doi: 10.1007/s00438-015-1130-7. Epub 2015 Oct 19 [PubMed PMID: 26481646]
Chen T, Tian L, Wang X, Fan D, Ma G, Tang R, Xuan X. Possible misdiagnosis of 46,XX testicular disorders of sex development in infertile males. International journal of medical sciences. 2020:17(9):1136-1141. doi: 10.7150/ijms.46058. Epub 2020 May 11 [PubMed PMID: 32547308]
Miles B, Tadi P. Genetics, Somatic Mutation. StatPearls. 2026 Jan:(): [PubMed PMID: 32491819]
Bin Shlhoob R, Tanaka J, Pandya AM. Genetics, X-Linked Inheritance. StatPearls. 2026 Jan:(): [PubMed PMID: 32491315]
Verma RS, Kleyman SM, Conte RA. Chromosomal mosaicisms during prenatal diagnosis. Gynecologic and obstetric investigation. 1998:45(1):12-5 [PubMed PMID: 9473156]
Lee HY, Koo DW, Lee JS. Lichen striatus colocalized with Becker's nevus: a case with two types of simultaneous cutaneous mosaicism? International journal of dermatology. 2020 Sep:59(9):e337-e339. doi: 10.1111/ijd.14921. Epub 2020 Jun 3 [PubMed PMID: 32492173]
Level 3 (low-level) evidenceBielanska M, Tan SL, Ao A. Chromosomal mosaicism throughout human preimplantation development in vitro: incidence, type, and relevance to embryo outcome. Human reproduction (Oxford, England). 2002 Feb:17(2):413-9 [PubMed PMID: 11821287]
Singla S, Iwamoto-Stohl LK, Zhu M, Zernicka-Goetz M. Autophagy-mediated apoptosis eliminates aneuploid cells in a mouse model of chromosome mosaicism. Nature communications. 2020 Jun 11:11(1):2958. doi: 10.1038/s41467-020-16796-3. Epub 2020 Jun 11 [PubMed PMID: 32528010]
Ge Y, Zhang J, Cai M, Chen X, Zhou Y. [Prenatal genetic analysis of three fetuses with abnormalities of chromosome 22]. Zhonghua yi xue yi chuan xue za zhi = Zhonghua yixue yichuanxue zazhi = Chinese journal of medical genetics. 2020 Apr 10:37(4):405-409. doi: 10.3760/cma.j.issn.1003-9406.2020.04.010. Epub [PubMed PMID: 32219823]
Taylor TH, Gitlin SA, Patrick JL, Crain JL, Wilson JM, Griffin DK. The origin, mechanisms, incidence and clinical consequences of chromosomal mosaicism in humans. Human reproduction update. 2014 Jul-Aug:20(4):571-81. doi: 10.1093/humupd/dmu016. Epub 2014 Mar 25 [PubMed PMID: 24667481]
Ramos C, Ocampos M, Barbato IT, Graça Bicalho MD, Nisihara R. Molecular analysis of FMR1 gene in a population in Southern Brazil: Comparison of four methods. Practical laboratory medicine. 2020 Aug:21():e00162. doi: 10.1016/j.plabm.2020.e00162. Epub 2020 May 6 [PubMed PMID: 32426440]
McCoy RC. Mosaicism in Preimplantation Human Embryos: When Chromosomal Abnormalities Are the Norm. Trends in genetics : TIG. 2017 Jul:33(7):448-463. doi: 10.1016/j.tig.2017.04.001. Epub 2017 Apr 28 [PubMed PMID: 28457629]
Dörnen J, Sieler M, Weiler J, Keil S, Dittmar T. Cell Fusion-Mediated Tissue Regeneration as an Inducer of Polyploidy and Aneuploidy. International journal of molecular sciences. 2020 Mar 6:21(5):. doi: 10.3390/ijms21051811. Epub 2020 Mar 6 [PubMed PMID: 32155721]
Jang J, Engleka KA, Liu F, Li L, Song G, Epstein JA, Li D. An Engineered Mouse to Identify Proliferating Cells and Their Derivatives. Frontiers in cell and developmental biology. 2020:8():388. doi: 10.3389/fcell.2020.00388. Epub 2020 May 25 [PubMed PMID: 32523954]
Campbell IM, Shaw CA, Stankiewicz P, Lupski JR. Somatic mosaicism: implications for disease and transmission genetics. Trends in genetics : TIG. 2015 Jul:31(7):382-92. doi: 10.1016/j.tig.2015.03.013. Epub 2015 Apr 21 [PubMed PMID: 25910407]
Zachaki S, Kouvidi E, Pantou A, Tsarouha H, Mitrakos A, Tounta G, Charalampous I, Manola KN, Kanavakis E, Mavrou A. Low-level X Chromosome Mosaicism: A Common Finding in Women Undergoing IVF. In vivo (Athens, Greece). 2020 May-Jun:34(3):1433-1437. doi: 10.21873/invivo.11925. Epub [PubMed PMID: 32354942]
Middelkamp S, van Tol HTA, Spierings DCJ, Boymans S, Guryev V, Roelen BAJ, Lansdorp PM, Cuppen E, Kuijk EW. Sperm DNA damage causes genomic instability in early embryonic development. Science advances. 2020 Apr:6(16):eaaz7602. doi: 10.1126/sciadv.aaz7602. Epub 2020 Apr 15 [PubMed PMID: 32494621]
Level 3 (low-level) evidenceNeofytou M. Predicting fetoplacental mosaicism during cfDNA-based NIPT. Current opinion in obstetrics & gynecology. 2020 Apr:32(2):152-158. doi: 10.1097/GCO.0000000000000610. Epub [PubMed PMID: 31977337]
Level 3 (low-level) evidenceBlakey-Cheung S, Parker P, Schlaff W, Monseur B, Keppler-Noreuil K, Al-Kouatly HB. Diagnosis and clinical delineation of mosaic tetrasomy 5p. European journal of medical genetics. 2020 Jan:63(1):103634. doi: 10.1016/j.ejmg.2019.02.006. Epub 2019 Feb 21 [PubMed PMID: 30797979]
Grønskov K, Rosenberg T, Sand A, Brøndum-Nielsen K. Mutational analysis of PAX6: 16 novel mutations including 5 missense mutations with a mild aniridia phenotype. European journal of human genetics : EJHG. 1999 Apr:7(3):274-86 [PubMed PMID: 10234503]
Dubey SK, Mahalaxmi N, Vijayalakshmi P, Sundaresan P. Mutational analysis and genotype-phenotype correlations in southern Indian patients with sporadic and familial aniridia. Molecular vision. 2015:21():88-97 [PubMed PMID: 25678763]
Level 2 (mid-level) evidenceCôté GB. The cis-trans effects of crossing-over on the penetrance and expressivity of dominantly inherited disorders. Annales de genetique. 1989:32(3):132-5 [PubMed PMID: 2817771]
Tesner P, Drabova J, Stolfa M, Kudr M, Kyncl M, Moslerova V, Novotna D, Kremlikova Pourova R, Kocarek E, Rasplickova T, Sedlacek Z, Vlckova M. A boy with developmental delay and mosaic supernumerary inv dup(5)(p15.33p15.1) leading to distal 5p tetrasomy - case report and review of the literature. Molecular cytogenetics. 2018:11():29. doi: 10.1186/s13039-018-0377-1. Epub 2018 May 9 [PubMed PMID: 29760779]
Level 3 (low-level) evidenceGaspar IM, Gaspar A. Variable expression and penetrance in Portuguese families with Familial Hypercholesterolemia with mild phenotype. Atherosclerosis. Supplements. 2019 Mar:36():28-30. doi: 10.1016/j.atherosclerosissup.2019.01.006. Epub [PubMed PMID: 30876530]
Fanarraga ML, Griffiths IR, McCulloch MC, Barrie JA, Cattanach BM, Brophy PJ, Kennedy PG. Rumpshaker: an X-linked mutation affecting CNS myelination. A study of the female heterozygote. Neuropathology and applied neurobiology. 1991 Aug:17(4):323-34 [PubMed PMID: 1944806]
Level 3 (low-level) evidenceCunha KS, Simioni M, Vieira TP, Gil-da-Silva-Lopes VL, Puzzi MB, Steiner CE. Tetrasomy 3q26.32-q29 due to a supernumerary marker chromosome in a child with pigmentary mosaicism of Ito. Genetics and molecular biology. 2016 Mar:39(1):35-9. doi: 10.1590/1678-4685-GMB-2015-0033. Epub [PubMed PMID: 27007896]
Waseem M, Mazhar M. Agammaglobulinemia. StatPearls. 2026 Jan:(): [PubMed PMID: 32310401]
Dhar SU, Robbins-Furman P, Levy ML, Patel A, Scaglia F. Tetrasomy 13q mosaicism associated with phylloid hypomelanosis and precocious puberty. American journal of medical genetics. Part A. 2009 May:149A(5):993-6. doi: 10.1002/ajmg.a.32758. Epub [PubMed PMID: 19334087]
Level 3 (low-level) evidenceSachdev NM, Maxwell SM, Besser AG, Grifo JA. Diagnosis and clinical management of embryonic mosaicism. Fertility and sterility. 2017 Jan:107(1):6-11. doi: 10.1016/j.fertnstert.2016.10.006. Epub 2016 Nov 11 [PubMed PMID: 27842993]
Guo X, Dai X, Zhou T, Wang H, Ni J, Xue J, Wang X. Mosaic loss of human Y chromosome: what, how and why. Human genetics. 2020 Apr:139(4):421-446. doi: 10.1007/s00439-020-02114-w. Epub 2020 Feb 4 [PubMed PMID: 32020362]
Zitzmann M, Rohayem J. Gonadal dysfunction and beyond: Clinical challenges in children, adolescents, and adults with 47,XXY Klinefelter syndrome. American journal of medical genetics. Part C, Seminars in medical genetics. 2020 Jun:184(2):302-312. doi: 10.1002/ajmg.c.31786. Epub 2020 May 16 [PubMed PMID: 32415901]
Rodríguez-Martín C, Robledo C, Gómez-Mariano G, Monzón S, Sastre A, Abelairas J, Sábado C, Martín-Begué N, Ferreres JC, Fernández-Teijeiro A, González-Campora R, Rios-Moreno MJ, Zaballos Á, Cuesta I, Martínez-Delgado B, Posada M, Alonso J. Frequency of low-level and high-level mosaicism in sporadic retinoblastoma: genotype-phenotype relationships. Journal of human genetics. 2020 Jan:65(2):165-174. doi: 10.1038/s10038-019-0696-z. Epub 2019 Nov 26 [PubMed PMID: 31772335]
Gonzalez Garcia A, Malone J, Li H. A novel mosaic variant on SMC1A reported in buccal mucosa cells, albeit not in blood, of a patient with Cornelia de Lange-like presentation. Cold Spring Harbor molecular case studies. 2020 Jun:6(3):. doi: 10.1101/mcs.a005322. Epub 2020 Jun 12 [PubMed PMID: 32532882]
Level 3 (low-level) evidenceTakeguchi R, Takahashi S, Kuroda M, Tanaka R, Suzuki N, Tomonoh Y, Ihara Y, Sugiyama N, Itoh M. MeCP2_e2 partially compensates for lack of MeCP2_e1: A male case of Rett syndrome. Molecular genetics & genomic medicine. 2020 Feb:8(2):e1088. doi: 10.1002/mgg3.1088. Epub 2019 Dec 9 [PubMed PMID: 31816669]
Level 3 (low-level) evidenceCohen AS, Wilson SL, Trinh J, Ye XC. Detecting somatic mosaicism: considerations and clinical implications. Clinical genetics. 2015 Jun:87(6):554-62. doi: 10.1111/cge.12502. Epub 2014 Oct 7 [PubMed PMID: 25223253]
Chauhan PR, Tanaka J. Genetics, Chromosome Abnormalities. StatPearls. 2026 Jan:(): [PubMed PMID: 32491623]
Grati FR, Malvestiti F, Branca L, Agrati C, Maggi F, Simoni G. Chromosomal mosaicism in the fetoplacental unit. Best practice & research. Clinical obstetrics & gynaecology. 2017 Jul:42():39-52. doi: 10.1016/j.bpobgyn.2017.02.004. Epub 2017 Feb 17 [PubMed PMID: 28284509]
Alfirevic Z, Navaratnam K, Mujezinovic F. Amniocentesis and chorionic villus sampling for prenatal diagnosis. The Cochrane database of systematic reviews. 2017 Sep 4:9(9):CD003252. doi: 10.1002/14651858.CD003252.pub2. Epub 2017 Sep 4 [PubMed PMID: 28869276]
Level 1 (high-level) evidenceTara F, Lotfalizadeh M, Moeindarbari S. The effect of diagnostic amniocentesis and its complications on early spontaneous abortion. Electronic physician. 2016 Aug:8(8):2787-2792 [PubMed PMID: 27757190]
Wapner RJ, Martin CL, Levy B, Ballif BC, Eng CM, Zachary JM, Savage M, Platt LD, Saltzman D, Grobman WA, Klugman S, Scholl T, Simpson JL, McCall K, Aggarwal VS, Bunke B, Nahum O, Patel A, Lamb AN, Thom EA, Beaudet AL, Ledbetter DH, Shaffer LG, Jackson L. Chromosomal microarray versus karyotyping for prenatal diagnosis. The New England journal of medicine. 2012 Dec 6:367(23):2175-84. doi: 10.1056/NEJMoa1203382. Epub [PubMed PMID: 23215555]
Iwata-Otsubo A, Radke B, Findley S, Abernathy B, Vallejos CE, Jackson SA. Fluorescence In Situ Hybridization (FISH)-Based Karyotyping Reveals Rapid Evolution of Centromeric and Subtelomeric Repeats in Common Bean (Phaseolus vulgaris) and Relatives. G3 (Bethesda, Md.). 2016 Apr 7:6(4):1013-22. doi: 10.1534/g3.115.024984. Epub 2016 Apr 7 [PubMed PMID: 26865698]
Findlay I, Corby N, Rutherford A, Quirke P. Comparison of FISH PRINS, and conventional and fluorescent PCR for single-cell sexing: suitability for preimplantation genetic diagnosis. Journal of assisted reproduction and genetics. 1998 May:15(5):258-65 [PubMed PMID: 9604757]
Notini AJ, Craig JM, White SJ. Copy number variation and mosaicism. Cytogenetic and genome research. 2008:123(1-4):270-7. doi: 10.1159/000184717. Epub 2009 Mar 11 [PubMed PMID: 19287164]
Shah MS, Cinnioglu C, Maisenbacher M, Comstock I, Kort J, Lathi RB. Comparison of cytogenetics and molecular karyotyping for chromosome testing of miscarriage specimens. Fertility and sterility. 2017 Apr:107(4):1028-1033. doi: 10.1016/j.fertnstert.2017.01.022. Epub 2017 Mar 7 [PubMed PMID: 28283267]
Cheng SSW, Chan KYK, Leung KKP, Au PKC, Tam WK, Li SKM, Luk HM, Kan ASY, Chung BHY, Lo IFM, Tang MHY. Experience of chromosomal microarray applied in prenatal and postnatal settings in Hong Kong. American journal of medical genetics. Part C, Seminars in medical genetics. 2019 Jun:181(2):196-207. doi: 10.1002/ajmg.c.31697. Epub 2019 Mar 23 [PubMed PMID: 30903683]
Shi Q, Qiu Y, Xu C, Yang H, Li C, Li N, Gao Y, Yu C. Next-generation sequencing analysis of each blastomere in good-quality embryos: insights into the origins and mechanisms of embryonic aneuploidy in cleavage-stage embryos. Journal of assisted reproduction and genetics. 2020 Jul:37(7):1711-1718. doi: 10.1007/s10815-020-01803-9. Epub 2020 May 22 [PubMed PMID: 32445153]
Level 2 (mid-level) evidencePaththinige CS, Sirisena ND, Kariyawasam UGIU, Dissanayake VHW. The Frequency and Spectrum of Chromosomal Translocations in a Cohort of Sri Lankans. BioMed research international. 2019:2019():9797104. doi: 10.1155/2019/9797104. Epub 2019 Apr 2 [PubMed PMID: 31061830]
Kaeser GE, Chun J. Mosaic Somatic Gene Recombination as a Potentially Unifying Hypothesis for Alzheimer's Disease. Frontiers in genetics. 2020:11():390. doi: 10.3389/fgene.2020.00390. Epub 2020 May 7 [PubMed PMID: 32457796]
Papavassiliou P, York TP, Gursoy N, Hill G, Nicely LV, Sundaram U, McClain A, Aggen SH, Eaves L, Riley B, Jackson-Cook C. The phenotype of persons having mosaicism for trisomy 21/Down syndrome reflects the percentage of trisomic cells present in different tissues. American journal of medical genetics. Part A. 2009 Feb 15:149A(4):573-83. doi: 10.1002/ajmg.a.32729. Epub [PubMed PMID: 19291777]
Armstrong JM, Malhotra NR, Lau GA. Seminoma In A Young Phenotypic Female With Turner Syndrome 45,XO/46,XY Mosaicism: A Case Report With Review Of The Literature. Urology. 2020 May:139():168-170. doi: 10.1016/j.urology.2020.01.031. Epub 2020 Feb 10 [PubMed PMID: 32057790]
Level 3 (low-level) evidenceCastori M, Tadini G. Discoveries and controversies in cutaneous mosaicism. Giornale italiano di dermatologia e venereologia : organo ufficiale, Societa italiana di dermatologia e sifilografia. 2016 Jun:151(3):251-65 [PubMed PMID: 27070303]
Lichtenstein AV. Genetic Mosaicism and Cancer: Cause and Effect. Cancer research. 2018 Mar 15:78(6):1375-1378. doi: 10.1158/0008-5472.CAN-17-2769. Epub 2018 Feb 22 [PubMed PMID: 29472519]
Cohen PR. Segmental neurofibromatosis and cancer: report of triple malignancy in a woman with mosaic Neurofibromatosis 1 and review of neoplasms in segmental neurofibromatosis. Dermatology online journal. 2016 Jul 15:22(7):. pii: 13030/qt66k5j4wt. Epub 2016 Jul 15 [PubMed PMID: 27617721]
Machiela MJ. Mosaicism, aging and cancer. Current opinion in oncology. 2019 Mar:31(2):108-113. doi: 10.1097/CCO.0000000000000500. Epub [PubMed PMID: 30585859]
Level 3 (low-level) evidenceIourov IY, Vorsanova SG, Yurov YB, Kutsev SI. Ontogenetic and Pathogenetic Views on Somatic Chromosomal Mosaicism. Genes. 2019 May 19:10(5):. doi: 10.3390/genes10050379. Epub 2019 May 19 [PubMed PMID: 31109140]
Jelsig AM, Bertelsen B, Forss I, Karstensen JG. Two cases of somatic STK11 mosaicism in Danish patients with Peutz-Jeghers syndrome. Familial cancer. 2021 Jan:20(1):55-59. doi: 10.1007/s10689-020-00191-4. Epub 2020 Jun 6 [PubMed PMID: 32504210]
Level 3 (low-level) evidenceTournier-Lasserve E. Molecular Genetic Screening of CCM Patients: An Overview. Methods in molecular biology (Clifton, N.J.). 2020:2152():49-57. doi: 10.1007/978-1-0716-0640-7_4. Epub [PubMed PMID: 32524543]
Level 3 (low-level) evidenceMuyas F, Zapata L, Guigó R, Ossowski S. The rate and spectrum of mosaic mutations during embryogenesis revealed by RNA sequencing of 49 tissues. Genome medicine. 2020 May 27:12(1):49. doi: 10.1186/s13073-020-00746-1. Epub 2020 May 27 [PubMed PMID: 32460841]
Sasani TA, Pedersen BS, Gao Z, Baird L, Przeworski M, Jorde LB, Quinlan AR. Large, three-generation human families reveal post-zygotic mosaicism and variability in germline mutation accumulation. eLife. 2019 Sep 24:8():. doi: 10.7554/eLife.46922. Epub 2019 Sep 24 [PubMed PMID: 31549960]
Navarro-Cobos MJ, Balaton BP, Brown CJ. Genes that escape from X-chromosome inactivation: Potential contributors to Klinefelter syndrome. American journal of medical genetics. Part C, Seminars in medical genetics. 2020 Jun:184(2):226-238. doi: 10.1002/ajmg.c.31800. Epub 2020 May 22 [PubMed PMID: 32441398]
Youssoufian H, Pyeritz RE. Mechanisms and consequences of somatic mosaicism in humans. Nature reviews. Genetics. 2002 Oct:3(10):748-58 [PubMed PMID: 12360233]
Hall JG. Review and hypotheses: somatic mosaicism: observations related to clinical genetics. American journal of human genetics. 1988 Oct:43(4):355-63 [PubMed PMID: 3052049]
Hook EB, Warburton D. Turner syndrome revisited: review of new data supports the hypothesis that all viable 45,X cases are cryptic mosaics with a rescue cell line, implying an origin by mitotic loss. Human genetics. 2014 Apr:133(4):417-24. doi: 10.1007/s00439-014-1420-x. Epub 2014 Jan 30 [PubMed PMID: 24477775]
Level 3 (low-level) evidenceHu Q, Chai H, Shu W, Li P. Human ring chromosome registry for cases in the Chinese population: re-emphasizing Cytogenomic and clinical heterogeneity and reviewing diagnostic and treatment strategies. Molecular cytogenetics. 2018:11():19. doi: 10.1186/s13039-018-0367-3. Epub 2018 Feb 27 [PubMed PMID: 29492108]
Level 3 (low-level) evidencevan Dyk E, Pretorius PJ. Point mutation instability (PIN) mutator phenotype as model for true back mutations seen in hereditary tyrosinemia type 1 - a hypothesis. Journal of inherited metabolic disease. 2012 May:35(3):407-11. doi: 10.1007/s10545-011-9401-x. Epub 2011 Oct 15 [PubMed PMID: 22002443]
Ellis NA, Ciocci S, German J. Back mutation can produce phenotype reversion in Bloom syndrome somatic cells. Human genetics. 2001 Feb:108(2):167-73 [PubMed PMID: 11281456]
Hirschhorn R. In vivo reversion to normal of inherited mutations in humans. Journal of medical genetics. 2003 Oct:40(10):721-8 [PubMed PMID: 14569115]
Xiao H, Zhang Z, Li T, Zhang Q, Guo Q, Wu D, Wang H, Zhang M, Gao Y, Liao S. [Germinal mosaicism for partial deletion of the Dystrophin gene in a family affected with Duchenne muscular dystrophy]. Zhonghua yi xue yi chuan xue za zhi = Zhonghua yixue yichuanxue zazhi = Chinese journal of medical genetics. 2019 Oct 10:36(10):1015-1018. doi: 10.3760/cma.j.issn.1003-9406.2019.10.016. Epub [PubMed PMID: 31598949]