 Apoptosis and Cell Death: Signaling in Health and Diseases
Apoptosis and Cell Death: Signaling in Health and Diseases
Introduction
Apoptosis is a tightly regulated form of programmed cell death essential for normal development and tissue maintenance. This process serves as a checkpoint that balances cellular proliferation by eliminating potentially harmful cells, especially those that have undergone or are likely to undergo pathogenic transformation. This process is critical during embryogenesis, immune system regulation, and the preservation of tissue homeostasis throughout adulthood.[1] Advances in molecular biology have established apoptosis as a genetically programmed, adenosine triphosphate (ATP)-dependent, enzyme-mediated mechanism that enables the orderly dismantling of cells without provoking significant inflammation or injury to surrounding tissues.[2]
At the cellular level, apoptosis is characterized by coordinated structural and biochemical alterations, including cytoskeletal breakdown, chromatin condensation, DNA fragmentation, and membrane blebbing. Proteases and endonucleases activated through tightly regulated signaling pathways mediate these processes. Apoptosis contributes to organismal survival by eliminating cells that are no longer functional or that jeopardize tissue integrity. However, dysregulation of apoptotic pathways can disrupt normal homeostasis and contribute to disease pathogenesis through either excessive cell loss or impaired cell death (see Image. Mechanism of Apoptosis).[3] The apoptotic process is primarily mediated by caspases, a family of cysteine proteases activated through intrinsic and extrinsic signaling pathways. The intrinsic and extrinsic pathways may function independently or converge to initiate a coordinated cascade of enzymatic events that result in cellular disassembly and phagocytic clearance. Understanding apoptotic mechanisms is essential for elucidating disease pathogenesis, because dysregulation of programmed cell death is associated with a broad range of conditions, including malignant neoplasms, autoimmune disorders, neurodegenerative diseases, and ischemic injury.[4]
Causes
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Causes
Results from current research suggest that apoptosis is a predominant mechanism of cell death. Major cell death mechanisms are summarized below:
- Apoptosis: The best-characterized form of cell death is executed by caspase-3 or caspase-7 following activation of upstream caspase-8 (the extrinsic or death receptor pathway) or caspase-9 (the intrinsic or mitochondrial pathway, involving cytochrome c release). Apoptosis is immunologically silent and nonlytic, with cellular remnants cleared by phagocytosis.[5][6][7]
- Necroptosis: This caspase-independent form of programmed necrosis is triggered by inhibition of caspase-8 via death receptor signaling (eg, tumor necrosis factor-α or Fas ligand). Receptor-interacting serine/threonine-protein kinase 3–dependent phosphorylation of mixed lineage kinase domain-like pseudokinase mediates necroptosis. Mixed lineage kinase domain-like pseudokinase then oligomerizes and disrupts the plasma membrane.[5][8][9]
- Pyroptosis: This inflammatory form of cell death is driven by inflammasome-activated caspase-1 (or caspase-4, caspase-5, or caspase-11), which cleaves gasdermin D. The N-terminal gasdermin D fragment forms pores in the plasma membrane, leading to cell lysis and the release of proinflammatory cytokines.[5][6][8][9][6]
- Ferroptosis: This iron-dependent, caspase-independent form of necrotic cell death is characterized by the accumulation of lethal lipid peroxides in cell membranes, typically resulting from failure of the glutathione peroxidase 4 antioxidant defense system.[10][11][12]
Although pyroptosis, apoptosis, and necroptosis were initially characterized as independent pathways, emerging studies have revealed significant molecular crosstalk among them. PANoptosis describes a distinct inflammatory cell death mechanism in which all 3 pathways are simultaneously engaged through multiprotein PANoptosome complexes. The resulting cell death cannot be fully accounted for by any single pathway alone.[13][14][15] Additionally, autophagy, primarily a prosurvival catabolic process that degrades cellular components via lysosomal machinery, can paradoxically contribute to cell death (autophagy-dependent cell death) when dysregulated or hyperactivated.[16][17][18] Finally, anastasis refers to the reversal of late-stage apoptosis, even after executioner caspase activation, and has been observed in cancer cell lines, where recovered cells may acquire increased invasiveness and genomic instability.[19][20][21]
Anatomical Pathology
Apoptosis is morphologically distinct from other forms of cell death and is identifiable by both light and electron microscopy. Apoptotic cells undergo shrinkage due to caspase-mediated cleavage of cytoskeletal proteins, resulting in the collapse of the cellular architecture. The cytoplasm becomes deeply eosinophilic because the cell condenses and separates from adjacent cells with loss of cell-to-cell contact.[22][23][24][25]
The morphologic hallmark of apoptosis is pyknosis, in which nuclear chromatin condenses into one or more basophilic masses against the nuclear envelope. Pyknosis is followed by karyorrhexis, the fragmentation of the condensed nucleus. Caspase-activated DNase (CAD) drives this process by cleaving DNA at internucleosomal sites, producing the characteristic DNA ladder pattern on gel electrophoresis.[22][26][27][28]
In the final stages, the condensed cytoplasm and fragmented nucleus break into membrane-bound fragments called apoptotic bodies, which bud from the cell surface. Notably, the plasma membrane remains intact throughout the process. Phosphatidylserine becomes externalized on the outer leaflet of the plasma membrane, signaling rapid recognition and engulfment by macrophages and neighboring cells in a process termed efferocytosis. Apoptosis remains immunologically silent before cellular contents are released into the extracellular space. Consequently, apoptosis does not provoke an inflammatory response, in contrast to necrosis, necroptosis, pyroptosis, and ferroptosis, all of which involve plasma membrane rupture, cell swelling, and release of proinflammatory intracellular contents.[24][29][30][31][32]
Clinical Pathology
Detecting apoptosis effectively requires multiple assays, because no single method can evaluate its multistage complexity. The following sections provide an overview of key assays organized by their specific detection targets.
Cytomorphological Changes
Apoptotic cells exhibit characteristic morphological features, including chromatin condensation, nuclear fragmentation, and plasma membrane blebbing. Chromatin condensation can be visualized using DNA-binding fluorescent dyes such as 4′,6-diamidino-2-phenylindole (DAPI) or Hoechst, which produce increased fluorescence intensity in condensed nuclei during microscopic assessment. Membrane blebbing can be observed in live cells using phase-contrast microscopy and is associated with caspase-mediated cleavage of cytoskeletal regulatory proteins, including gelsolin and Rho-associated coiled-coil-containing protein kinase 1 (ROCK1). Although fixed-cell analysis using caspase substrate markers can aid in detecting apoptosis, this method may generate false-positive results due to nonspecific staining or late-stage cellular degradation.[3]
An alternative method for assessing apoptosis is annexin V staining combined with propidium iodide or 7-aminoactinomycin D (7-AAD) in flow cytometry. Annexin V binds phosphatidylserine residues that become externalized on the outer leaflet of the plasma membrane early in apoptosis. Notably, propidium iodide or 7-AAD distinguishes late apoptotic or necrotic cells by labeling cells with compromised membrane integrity. This approach enables quantitative discrimination among viable, early apoptotic, late apoptotic, and necrotic cell populations, providing greater sensitivity and specificity than morphology-based assessment alone.[33]
DNA Fragmentation
A hallmark of late-stage apoptosis is the cleavage of DNA into 180 to 200 base-pair fragments. The terminal deoxynucleotidyl transferase deoxyuridine triphosphate nick-end labeling (TUNEL) assay detects DNA fragmentation by labeling the exposed 3′-OH termini of DNA strand breaks with fluorescent or chromogenic nucleotides, enabling visualization with fluorescence microscopy or quantification with flow cytometry. Alternatively, DNA laddering analysis using agarose gel electrophoresis provides a cost-effective method for detecting apoptotic DNA fragmentation. However, DNA laddering analysis is generally less sensitive and less quantitative than TUNEL-based approaches. TUNEL positivity should be interpreted cautiously, because DNA strand breaks may also occur in other forms of cell death or during DNA repair processes.[34][35]
Caspase Activity, Cleaved Substrates, and Regulatory Proteins
Caspase activation, particularly of caspases-3, -8, and -9, can be measured using fluorogenic substrates or fluorescent inhibitors. Midstage apoptosis is often confirmed by detecting cleavage of poly(ADP-ribose) polymerase via Western blot analysis. Additionally, the balance between proapoptotic and antiapoptotic proteins, such as BCL-2 and BAX, can be assessed using immunostaining assays to evaluate regulatory mechanisms.[36]
Membrane Changes
Early apoptotic stages are characterized by phosphatidylserine externalization, detected by annexin V staining, typically combined with propidium iodide to distinguish apoptotic from necrotic cells. In later stages, loss of membrane integrity is monitored using dye-exclusion assays.[37]
Whole-Mount Detection
Apoptosis in embryos or tissues can be detected using acidophilic dyes such as acridine orange, Nile blue sulfate, and neutral red. However, these dyes require validation due to nonspecific staining of debris and have limitations, including toxicity and poor penetration. LysoTracker Red enables stable, 3D imaging in thick tissues via confocal microscopy.[38]
Mitochondrial Assays
Mitochondrial assays for apoptosis detection focus on key changes, including loss of mitochondrial membrane potential, detected using dyes such as JC-1, tetramethylrhodamine methyl ester, or tetramethylrhodamine ethyl ester.[3] Permeability transition pore opening is monitored by quenching of calcein fluorescence. In addition, cytochrome c release is tracked by immunofluorescence or subcellular fractionation. These assays are often combined with caspase activity measurements to improve specificity.[39]
Mechanisms
Cellular proliferation and cell death are balanced in all normal tissues of multicellular organisms. Physiological cell death is vital for cellular development and function. This process is referred to as apoptosis and involves several pathways.
Intrinsic Pathway
The intrinsic pathway is activated when cells are exposed to internal or external stressors, including DNA damage from ionizing radiation, UV light, or chemotherapeutic agents, hypoxia, or the accumulation of misfolded proteins within the cell, as occurs in conditions such as Alzheimer disease, Parkinson disease, or Huntington disease. When a cell undergoes stress, cytochrome c leaks from the mitochondrial intermembrane space into the cytosol, leading to the activation of caspase-9. The BCL2 and TP53 genes are essential regulators of this pathway.[36][37]
The BCL-2 protein family includes both proapoptotic and antiapoptotic members. Proapoptotic proteins detect death signals and trigger cell death, whereas antiapoptotic proteins block this process. These evolutionarily conserved proteins can act through diverse mechanisms across species. Different cellular signals can alter their activity and location within the cell, creating a complex network that balances cell survival and death. Because of their key role in regulating apoptosis, BCL-2 family proteins play a pivotal role in embryonic development and adult homeostasis.[39][40]
Extrinsic Pathway
The extrinsic pathway is activated when a cell receives death signals from neighboring cells or immune cells. This receptor-mediated pathway is initiated by ligands that bind to death receptors on the cell surface, thereby triggering apoptosis. The process relies on the interaction between cell-surface death receptors and their corresponding ligands, ultimately leading to the activation of caspase-8, a key initiator of the apoptotic cascade.
- Tumor necrosis factor-α (TNF-α): A cytokine produced by macrophages and is the major extrinsic mediator of apoptosis. TNF-α binds to the tumor necrosis factor receptor (TNFR1), thereby activating caspases.
- Fas: Fas is a transmembrane death receptor of the tumor necrosis factor family that is expressed on the surface of T cells. Fas expression increases during infection. Following activation, T lymphocytes release Fas ligand, which binds to Fas on the same or neighboring cells, initiating caspase activation and subsequent apoptosis. This mechanism plays a critical role in immune homeostasis by eliminating activated T lymphocytes once the infection has resolved.
- BCL2 genes: The antiapoptotic BCL2 genes are located on chromosome 18 and encode the BCL-2 protein. BCL-2 inhibits apoptotic signaling and prevents the release of cytochrome c from the mitochondria. Cytochrome c is located between the inner and outer mitochondrial membranes. Once released, cytochrome c can bind to apoptotic protease-activating factor 1 (APAF1), activating procaspase-9.
- TP53: TP53 is a tumor suppressor gene that encodes the p53 protein, which regulates the cell cycle and promotes tumor suppression. Following DNA damage, p53 arrests the cell cycle in the G1 phase, preventing the proliferation of cells with damaged DNA and facilitating DNA repair. Severe DNA damage induces apoptosis by activating the proapoptotic BAX gene. BAX proteins inhibit the antiapoptotic BCL-2 protein. The balance between proapoptotic and antiapoptotic genes regulates this process (see Image. DNA Repair and Apoptosis).
- Cytotoxic CD8+ T-cell pathway: CD8+ T cells secrete perforin, which forms pores in the target cells. Subsequently, these T cells secrete granzymes that enter target cells through these pores, activating caspases.
- Caspases: Caspases are a group of proteases expressed in cells in an inactive form that require proteolytic cleavage for activation. These enzymes are the primary effectors of apoptotic responses and are activated by several regulators.[41]
- Initiator caspases: Initiator caspases include caspase-2, caspase-8, caspase-9, and caspase-10. When activated, these enzymes promote the activation of effector caspases.
- Effector caspases: Effector caspases include caspase-3, caspase-6, and caspase-7. Active effector caspases cleave several cellular proteins, leading to cell death, phagocytosis, and removal of cellular debris.
- Caspase-3: Caspase-3 is the most frequently activated caspase and catalyzes the cleavage of major cellular proteins and the condensation of chromatin. This enzyme also activates deoxyribonucleases, which cause DNA fragmentation followed by internucleosomal fragmentation.[42][43]
Execution Pathway
The execution phase is the final irreversible step of apoptosis, during which effector caspases, including caspase-3, caspase-6, and caspase-7, break down the cell. Upstream caspases from the extrinsic or intrinsic pathway activate these effector caspases. The enzymes degrade the nuclear envelope, block DNA repair, and fragment DNA into a distinct pattern via an activated nuclease, thereby marking cells for irreversible breakdown. The cell shrinks, alters its membrane to expose phagocytic signals, and forms debris-filled vesicles, which are cleared by phagocytes to prevent inflammation. Proapoptotic and antiapoptotic proteins tightly regulate this orderly process.[44]
Other Players
Following initial cell death, danger-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs) are released from apoptotic cells, stimulating the release of additional inflammatory mediators. The extent of this inflammatory response depends on the type of cell death and the involvement of other signaling mechanisms. Consequently, the extent to which apoptosis is immunologically silent remains incompletely understood. In several pathological conditions, such as cancer, apoptotic pathways are dysregulated by inhibitors of apoptosis proteins (IAPs), a family of antiapoptotic modulators that suppress programmed cell death.[45]
Clinicopathologic Correlations
Embryogenesis
During embryonic development, apoptosis of interdigital tissue precedes digit formation. Similarly, several body cavities undergo apoptosis in utero.[46] For example, during male fetal development, Müllerian structures regress due to Müllerian inhibiting substance synthesized by Sertoli cells.[47]
Menstrual Cycle
The cyclical shedding of the endometrium following the withdrawal of estrogen and progesterone during the late luteal phase of the menstrual cycle represents a physiological process mediated by apoptosis, ischemic necrosis, and an inflammatory cascade secondary to spiral arteriole vasoconstriction.[47]
Immunological Regulation
- Virus-infected cells: Cytotoxic T cells eliminate virus-infected cells by apoptosis.
- Cells with DNA damage: Cells with DNA damage caused by radiation exposure or chemotherapeutic agents are arrested in the G1 phase of the cell cycle to facilitate repair through activation of p53. TP53 is a tumor suppressor gene. A TP53 mutation inhibits apoptosis, leading to the survival of abnormal cells and the development of carcinomas.
- Autoreactive T cells: Autoreactive T cells in the thymus are eliminated through apoptosis.[48]
Apoptosis is required for the development and maintenance of the immune system. Newly formed B and T lymphocytes undergo a checkpoint to assess their response to self-antigens. Autoreactive cells are eliminated by apoptosis to prevent autoimmune disease. Apoptosis is also necessary to terminate the immune response after a pathogen is cleared from the body, including by removing acute inflammatory cells, such as neutrophils, from healing sites. Furthermore, corticosteroid-induced lymphocytolysis of B and T lymphocytes is mediated through apoptotic pathways.[24]
Removal of Misfolded Proteins
Apoptosis facilitates the clearance of misfolded proteins, including amyloid aggregates and prion-associated proteins. Consequently, this mechanism mitigates the accumulation of pathological protein aggregates implicated in the pathogenesis of neurodegenerative diseases, such as Alzheimer disease.
Clinical Significance
Dysregulation of apoptosis is a hallmark of cancer and contributes to the pathogenesis of autoimmune, neurodegenerative, and cardiovascular diseases.[49][50][51] The clinical implications of aberrant apoptotic signaling span virtually every organ system and have driven transformative advances in targeted therapeutics.
Tumorigenesis: Evasion of apoptosis is a defining feature of malignant transformation. In follicular lymphoma, the t(14;18) chromosomal translocation juxtaposes the BCL2 gene with the immunoglobulin heavy chain locus, resulting in constitutive overexpression of the antiapoptotic BCL-2 protein, which prevents mitochondrial outer membrane permeabilization (MOMP), cytochrome c release, and caspase activation.[52] Loss-of-function mutations in TP53 impair transcriptional activation of proapoptotic BCL-2 homology 3 (BH3)-onlyproteins (PUMA, NOXA, BAX), allowing cells with unrepaired DNA damage to proliferate uncontrollably.[53] Germline TP53 mutations underlie Li-Fraumeni syndrome, conferring a markedly elevated lifetime risk of diverse malignant neoplasms. Environmental stressors such as exposure to chemical carcinogens, ionizing radiation, and oncogenic viruses can further affect p53 expression and function, enabling cells to bypass apoptotic checkpoints and accumulate additional oncogenic mutations.[49][53]
Autoimmune diseases: Defective apoptosis of autoreactive lymphocytes disrupts peripheral tolerance and contributes to autoimmune disorders, including systemic lupus erythematosus, rheumatoid arthritis, and autoimmune lymphoproliferative syndrome.[51][54] In systemic lupus erythematosus, impaired clearance of apoptotic debris exposes intracellular autoantigens (eg, histones and double-stranded DNA) to the immune system, driving pathogenic autoantibody production. Mitochondria-mediated a apoptotic pathways have also been implicated in the autoimmune destruction of pancreatic β-cells in type 1 diabetes.[51]
Neurodegenerative diseases: Excessive neuronal apoptosis is a central pathologic feature of Alzheimer disease, Parkinson disease, Huntington disease, and amyotrophic lateral sclerosis.[55][56] In Alzheimer disease, amyloid-β accumulation and tau hyperphosphorylation trigger mitochondrial dysfunction, oxidative stress, and caspase activation, leading to neuronal loss in the hippocampus and cerebral cortex.[57] In acute ischemic stroke, both necrosis and apoptosis contribute to neuronal death, with apoptosis predominating in the ischemic penumbra.[55]
Cardiovascular diseases: Although necrosis has been considered the sole mechanism of cardiomyocyte death in myocardial infarction, apoptosis has recently been recognized as a significant contributor, particularly during the reperfusion phase.[58][59] Reactive oxygen species generated during reperfusion activate both intrinsic (mitochondrial) and extrinsic (death receptor) apoptotic cascades, exacerbating myocardial injury beyond the initial ischemic insult.[59] Additionally, in atherosclerosis, apoptotic macrophages within advanced plaques promote necrotic core expansion and plaque instability, increasing the risk of rupture and acute coronary events.[60]
Therapeutic implications: Advances in the molecular characterization of apoptotic signaling pathways have identified clinically actionable therapeutic targets. Venetoclax, a selective BCL-2 inhibitor and the first US Food and Drug Administrationapproved BH3 mimetic, has transformed the treatment of chronic lymphocytic leukemia and acute myeloid leukemia, with next-generation agents (sonrotoclax, lisaftoclax) and inhibitors of MCL-1 and BCL-XL in clinical development.[61][62][63] Antibody-drug conjugates (ADCs), such as trastuzumab emtansine and trastuzumab deruxtecan, exploit receptor-mediated endocytosis to deliver cytotoxic payloads that induce tumor cell apoptosis with enhanced specificity. Combinations of ADCs with immune checkpoint inhibitors have demonstrated synergistic antitumor activity.[64][65]
The TUNEL assay remains a widely used method for quantifying apoptotic cell death in pathologic tissue specimens. However, its inability to distinguish apoptosis from necrosis necessitates correlation with morphologic criteria and complementary assays.[66][67] Emerging strategies, including proteolysis-targeting chimeras (PROTACs) directed against antiapoptotic BCL-2 family members and dual-pathway inhibition approaches, represent the next frontier in precision oncology aimed at restoring apoptotic competence in treatment-resistant malignant neoplasms.[63][68]
Media
(Click Image to Enlarge)
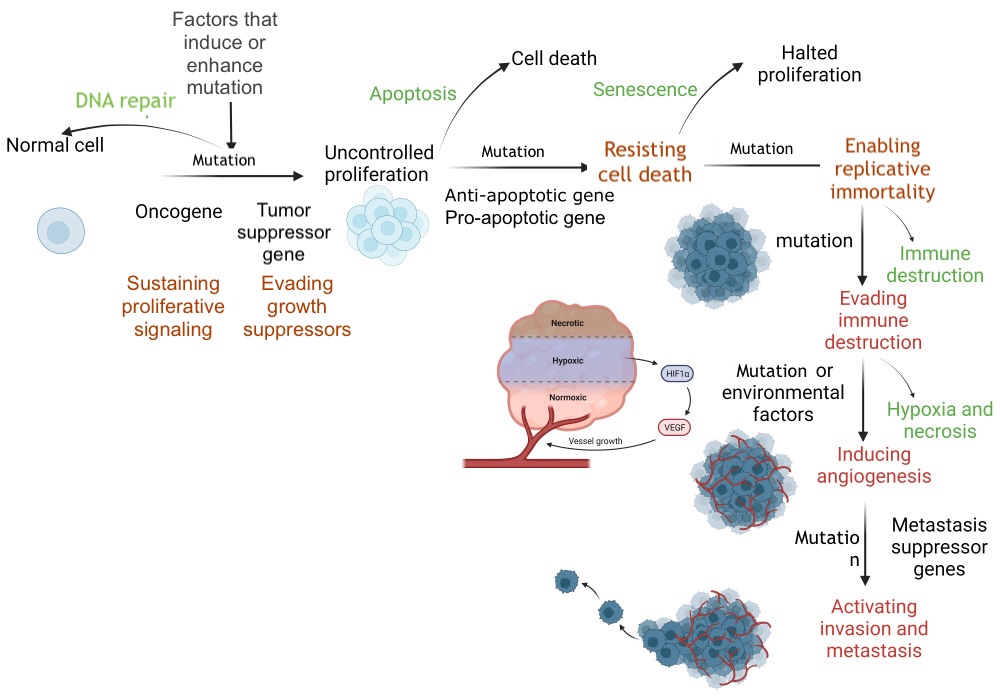
DNA Repair and Apoptosis. DNA repair restores a cell to its normal state, whereas apoptosis eliminates a cell without inflammation. Defective DNA repair and apoptosis allow the cell to initiate uncontrolled proliferation through promotion and subsequent progression, resulting in distant tissue invasion. HIF-1α, hypoxia-inducible factor 1 alpha; VEGF, vascular endothelial growth factor.
Contributed by S Abd El Fattah, MD
(Click Image to Enlarge)
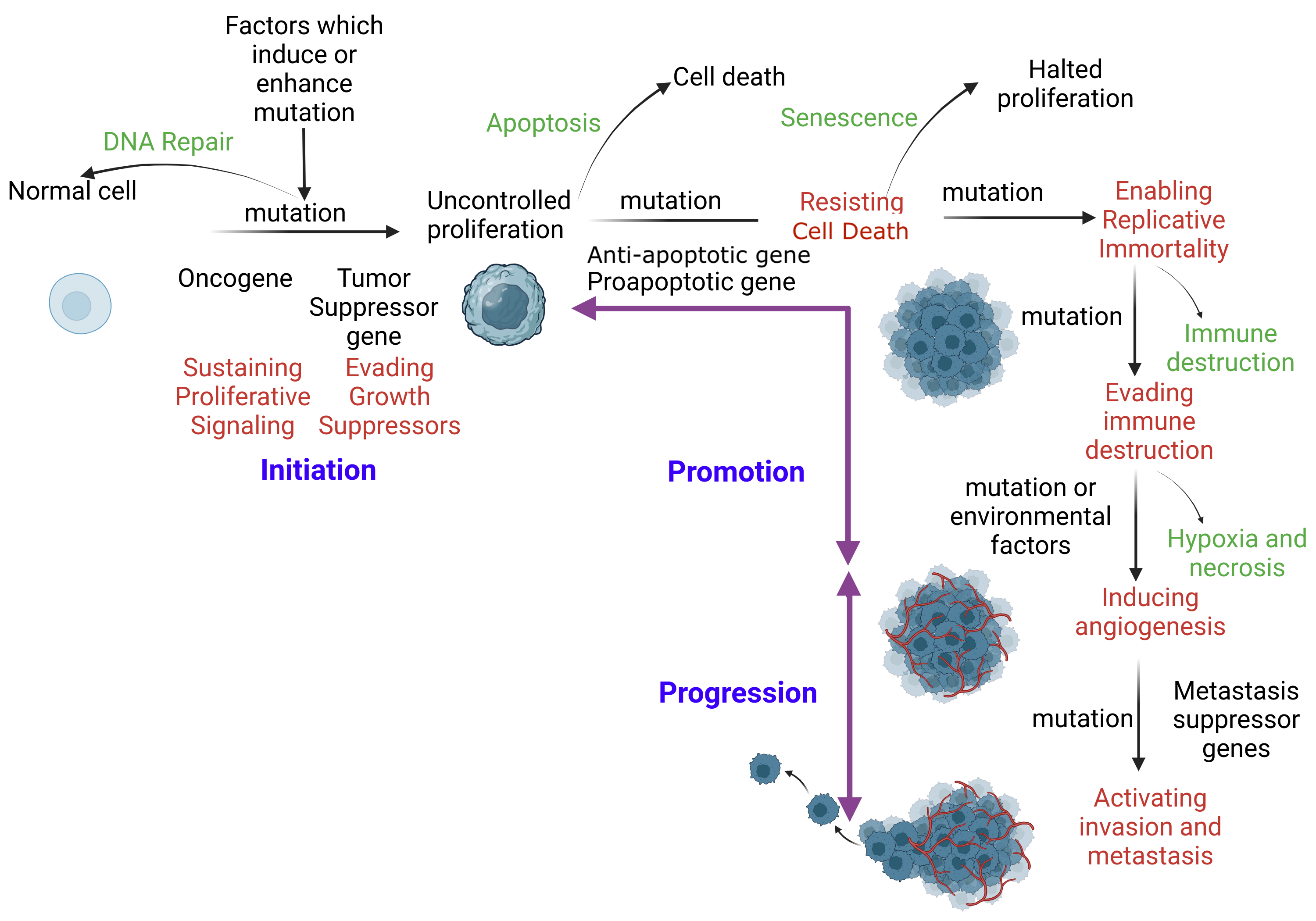
Mechanism of Apoptosis. When DNA repair fails, apoptosis ensues to eliminate the corrupt cell without inflammation. An initiated cell evades tumor suppressor genes and activates constitutional proliferation (auto-proliferation). Subsequently, the initiated cell is promoted through the acquisition of additional mutations. This mutation immortalizes the cell by skipping immune checkpoints. Additional mutations are acquired, facilitating distant tissue invasion.
SA Ibrahim, MBBCh, MSc, PhD
References
Park W, Wei S, Kim BS, Kim B, Bae SJ, Chae YC, Ryu D, Ha KT. Diversity and complexity of cell death: a historical review. Experimental & molecular medicine. 2023 Aug:55(8):1573-1594. doi: 10.1038/s12276-023-01078-x. Epub 2023 Aug 23 [PubMed PMID: 37612413]
Galluzzi L, Vitale I, Aaronson SA, Abrams JM, Adam D, Agostinis P, Alnemri ES, Altucci L, Amelio I, Andrews DW, Annicchiarico-Petruzzelli M, Antonov AV, Arama E, Baehrecke EH, Barlev NA, Bazan NG, Bernassola F, Bertrand MJM, Bianchi K, Blagosklonny MV, Blomgren K, Borner C, Boya P, Brenner C, Campanella M, Candi E, Carmona-Gutierrez D, Cecconi F, Chan FK, Chandel NS, Cheng EH, Chipuk JE, Cidlowski JA, Ciechanover A, Cohen GM, Conrad M, Cubillos-Ruiz JR, Czabotar PE, D'Angiolella V, Dawson TM, Dawson VL, De Laurenzi V, De Maria R, Debatin KM, DeBerardinis RJ, Deshmukh M, Di Daniele N, Di Virgilio F, Dixit VM, Dixon SJ, Duckett CS, Dynlacht BD, El-Deiry WS, Elrod JW, Fimia GM, Fulda S, García-Sáez AJ, Garg AD, Garrido C, Gavathiotis E, Golstein P, Gottlieb E, Green DR, Greene LA, Gronemeyer H, Gross A, Hajnoczky G, Hardwick JM, Harris IS, Hengartner MO, Hetz C, Ichijo H, Jäättelä M, Joseph B, Jost PJ, Juin PP, Kaiser WJ, Karin M, Kaufmann T, Kepp O, Kimchi A, Kitsis RN, Klionsky DJ, Knight RA, Kumar S, Lee SW, Lemasters JJ, Levine B, Linkermann A, Lipton SA, Lockshin RA, López-Otín C, Lowe SW, Luedde T, Lugli E, MacFarlane M, Madeo F, Malewicz M, Malorni W, Manic G, Marine JC, Martin SJ, Martinou JC, Medema JP, Mehlen P, Meier P, Melino S, Miao EA, Molkentin JD, Moll UM, Muñoz-Pinedo C, Nagata S, Nuñez G, Oberst A, Oren M, Overholtzer M, Pagano M, Panaretakis T, Pasparakis M, Penninger JM, Pereira DM, Pervaiz S, Peter ME, Piacentini M, Pinton P, Prehn JHM, Puthalakath H, Rabinovich GA, Rehm M, Rizzuto R, Rodrigues CMP, Rubinsztein DC, Rudel T, Ryan KM, Sayan E, Scorrano L, Shao F, Shi Y, Silke J, Simon HU, Sistigu A, Stockwell BR, Strasser A, Szabadkai G, Tait SWG, Tang D, Tavernarakis N, Thorburn A, Tsujimoto Y, Turk B, Vanden Berghe T, Vandenabeele P, Vander Heiden MG, Villunger A, Virgin HW, Vousden KH, Vucic D, Wagner EF, Walczak H, Wallach D, Wang Y, Wells JA, Wood W, Yuan J, Zakeri Z, Zhivotovsky B, Zitvogel L, Melino G, Kroemer G. Molecular mechanisms of cell death: recommendations of the Nomenclature Committee on Cell Death 2018. Cell death and differentiation. 2018 Mar:25(3):486-541. doi: 10.1038/s41418-017-0012-4. Epub 2018 Jan 23 [PubMed PMID: 29362479]
Elmore S. Apoptosis: a review of programmed cell death. Toxicologic pathology. 2007 Jun:35(4):495-516 [PubMed PMID: 17562483]
Level 3 (low-level) evidenceKumar S, Dorstyn L, Lim Y. The role of caspases as executioners of apoptosis. Biochemical Society transactions. 2022 Feb 28:50(1):33-45. doi: 10.1042/BST20210751. Epub [PubMed PMID: 34940803]
Bertheloot D, Latz E, Franklin BS. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death. Cellular & molecular immunology. 2021 May:18(5):1106-1121. doi: 10.1038/s41423-020-00630-3. Epub 2021 Mar 30 [PubMed PMID: 33785842]
Ketelut-Carneiro N, Fitzgerald KA. Apoptosis, Pyroptosis, and Necroptosis-Oh My! The Many Ways a Cell Can Die. Journal of molecular biology. 2022 Feb 28:434(4):167378. doi: 10.1016/j.jmb.2021.167378. Epub 2021 Nov 25 [PubMed PMID: 34838807]
Ahmed A, Tait SWG. Molecular Cell Biology of Apoptosis in Health and Disease. Advances in experimental medicine and biology. 2025:1481():1-28. doi: 10.1007/978-3-031-92785-0_1. Epub [PubMed PMID: 41004078]
Level 3 (low-level) evidenceAi Y, Meng Y, Yan B, Zhou Q, Wang X. The biochemical pathways of apoptotic, necroptotic, pyroptotic, and ferroptotic cell death. Molecular cell. 2024 Jan 4:84(1):170-179. doi: 10.1016/j.molcel.2023.11.040. Epub [PubMed PMID: 38181758]
Yuan J, Ofengeim D. A guide to cell death pathways. Nature reviews. Molecular cell biology. 2024 May:25(5):379-395. doi: 10.1038/s41580-023-00689-6. Epub 2023 Dec 18 [PubMed PMID: 38110635]
Samir P, Malireddi RKS, Kanneganti TD. The PANoptosome: A Deadly Protein Complex Driving Pyroptosis, Apoptosis, and Necroptosis (PANoptosis). Frontiers in cellular and infection microbiology. 2020:10():238. doi: 10.3389/fcimb.2020.00238. Epub 2020 Jun 3 [PubMed PMID: 32582562]
Yan HF, Zou T, Tuo QZ, Xu S, Li H, Belaidi AA, Lei P. Ferroptosis: mechanisms and links with diseases. Signal transduction and targeted therapy. 2021 Feb 3:6(1):49. doi: 10.1038/s41392-020-00428-9. Epub 2021 Feb 3 [PubMed PMID: 33536413]
Fearnhead HO, Vandenabeele P, Vanden Berghe T. How do we fit ferroptosis in the family of regulated cell death? Cell death and differentiation. 2017 Dec:24(12):1991-1998. doi: 10.1038/cdd.2017.149. Epub 2017 Oct 6 [PubMed PMID: 28984871]
Sun X, Yang Y, Meng X, Li J, Liu X, Liu H. PANoptosis: Mechanisms, biology, and role in disease. Immunological reviews. 2024 Jan:321(1):246-262. doi: 10.1111/imr.13279. Epub 2023 Oct 12 [PubMed PMID: 37823450]
Chen W, Gullett JM, Tweedell RE, Kanneganti TD. Innate immune inflammatory cell death: PANoptosis and PANoptosomes in host defense and disease. European journal of immunology. 2023 Nov:53(11):e2250235. doi: 10.1002/eji.202250235. Epub 2023 Mar 3 [PubMed PMID: 36782083]
Qi Z, Zhu L, Wang K, Wang N. PANoptosis: Emerging mechanisms and disease implications. Life sciences. 2023 Nov 15:333():122158. doi: 10.1016/j.lfs.2023.122158. Epub 2023 Oct 6 [PubMed PMID: 37806654]
Level 2 (mid-level) evidenceHuang X, Yan H, Xu Z, Yang B, Luo P, He Q. The inducible role of autophagy in cell death: emerging evidence and future perspectives. Cell communication and signaling : CCS. 2025 Mar 26:23(1):151. doi: 10.1186/s12964-025-02135-w. Epub 2025 Mar 26 [PubMed PMID: 40140912]
Level 3 (low-level) evidenceBialik S, Dasari SK, Kimchi A. Autophagy-dependent cell death - where, how and why a cell eats itself to death. Journal of cell science. 2018 Sep 20:131(18):. pii: jcs215152. doi: 10.1242/jcs.215152. Epub 2018 Sep 20 [PubMed PMID: 30237248]
Denton D, Kumar S. Autophagy-dependent cell death. Cell death and differentiation. 2019 Mar:26(4):605-616. doi: 10.1038/s41418-018-0252-y. Epub 2018 Dec 19 [PubMed PMID: 30568239]
Chakraborty S, Mir KB, Seligson ND, Nayak D, Kumar R, Goswami A. Integration of EMT and cellular survival instincts in reprogramming of programmed cell death to anastasis. Cancer metastasis reviews. 2020 Jun:39(2):553-566. doi: 10.1007/s10555-020-09866-x. Epub [PubMed PMID: 32020420]
Sun L, Yao C, Li X, Wang Y, Wang R, Wang M, Liu Q, Montell DJ, Shao C, Gong Y, Sun G. Anastasis confers ovarian cancer cells increased malignancy through elevated p38 MAPK activation. Cell death and differentiation. 2023 Mar:30(3):809-824. doi: 10.1038/s41418-022-01081-1. Epub 2022 Nov 29 [PubMed PMID: 36447048]
Tang HL, Tang HM, Mak KH, Hu S, Wang SS, Wong KM, Wong CS, Wu HY, Law HT, Liu K, Talbot CC Jr, Lau WK, Montell DJ, Fung MC. Cell survival, DNA damage, and oncogenic transformation after a transient and reversible apoptotic response. Molecular biology of the cell. 2012 Jun:23(12):2240-52. doi: 10.1091/mbc.E11-11-0926. Epub 2012 Apr 25 [PubMed PMID: 22535522]
Doonan F, Cotter TG. Morphological assessment of apoptosis. Methods (San Diego, Calif.). 2008 Mar:44(3):200-4. doi: 10.1016/j.ymeth.2007.11.006. Epub [PubMed PMID: 18314050]
Hotchkiss RS, Strasser A, McDunn JE, Swanson PE. Cell death. The New England journal of medicine. 2009 Oct 15:361(16):1570-83. doi: 10.1056/NEJMra0901217. Epub [PubMed PMID: 19828534]
Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. British journal of cancer. 1972 Aug:26(4):239-57 [PubMed PMID: 4561027]
Level 3 (low-level) evidenceNdozangue-Touriguine O, Hamelin J, Bréard J. Cytoskeleton and apoptosis. Biochemical pharmacology. 2008 Jul 1:76(1):11-8. doi: 10.1016/j.bcp.2008.03.016. Epub 2008 Apr 1 [PubMed PMID: 18462707]
Martelli AM, Zweyer M, Ochs RL, Tazzari PL, Tabellini G, Narducci P, Bortul R. Nuclear apoptotic changes: an overview. Journal of cellular biochemistry. 2001:82(4):634-46 [PubMed PMID: 11500941]
Level 3 (low-level) evidenceSaraste A, Pulkki K. Morphologic and biochemical hallmarks of apoptosis. Cardiovascular research. 2000 Feb:45(3):528-37 [PubMed PMID: 10728374]
Vermeulen K, Van Bockstaele DR, Berneman ZN. Apoptosis: mechanisms and relevance in cancer. Annals of hematology. 2005 Oct:84(10):627-39 [PubMed PMID: 16041532]
Atkin-Smith GK, Poon IKH. Disassembly of the Dying: Mechanisms and Functions. Trends in cell biology. 2017 Feb:27(2):151-162. doi: 10.1016/j.tcb.2016.08.011. Epub 2016 Sep 16 [PubMed PMID: 27647018]
Yin C, Heit B. Cellular Responses to the Efferocytosis of Apoptotic Cells. Frontiers in immunology. 2021:12():631714. doi: 10.3389/fimmu.2021.631714. Epub 2021 Apr 20 [PubMed PMID: 33959122]
Schilperoort M, Ngai D, Sukka SR, Avrampou K, Shi H, Tabas I. The role of efferocytosis-fueled macrophage metabolism in the resolution of inflammation. Immunological reviews. 2023 Oct:319(1):65-80. doi: 10.1111/imr.13214. Epub 2023 May 9 [PubMed PMID: 37158427]
Doran AC, Yurdagul A Jr, Tabas I. Efferocytosis in health and disease. Nature reviews. Immunology. 2020 Apr:20(4):254-267. doi: 10.1038/s41577-019-0240-6. Epub 2019 Dec 10 [PubMed PMID: 31822793]
Crowley LC, Marfell BJ, Scott AP, Waterhouse NJ. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harbor protocols. 2016 Nov 1:2016(11):. doi: 10.1101/pdb.prot087288. Epub 2016 Nov 1 [PubMed PMID: 27803250]
Crowley LC, Marfell BJ, Waterhouse NJ. Detection of DNA Fragmentation in Apoptotic Cells by TUNEL. Cold Spring Harbor protocols. 2016 Oct 3:2016(10):. doi: 10.1101/pdb.prot087221. Epub 2016 Oct 3 [PubMed PMID: 27698233]
Moore CL, Savenka AV, Basnakian AG. TUNEL Assay: A Powerful Tool for Kidney Injury Evaluation. International journal of molecular sciences. 2021 Jan 2:22(1):. doi: 10.3390/ijms22010412. Epub 2021 Jan 2 [PubMed PMID: 33401733]
Lossi L. The concept of intrinsic versus extrinsic apoptosis. The Biochemical journal. 2022 Feb 11:479(3):357-384. doi: 10.1042/BCJ20210854. Epub [PubMed PMID: 35147165]
Kashyap D, Garg VK, Goel N. Intrinsic and extrinsic pathways of apoptosis: Role in cancer development and prognosis. Advances in protein chemistry and structural biology. 2021:125():73-120. doi: 10.1016/bs.apcsb.2021.01.003. Epub 2021 Apr 15 [PubMed PMID: 33931145]
Level 3 (low-level) evidenceWoods CG, Heuvel JP, Rusyn I. Genomic profiling in nuclear receptor-mediated toxicity. Toxicologic pathology. 2007 Jun:35(4):474-94 [PubMed PMID: 17562482]
Warren CFA, Wong-Brown MW, Bowden NA. BCL-2 family isoforms in apoptosis and cancer. Cell death & disease. 2019 Feb 21:10(3):177. doi: 10.1038/s41419-019-1407-6. Epub 2019 Feb 21 [PubMed PMID: 30792387]
Chan SL, Yu VC. Proteins of the bcl-2 family in apoptosis signalling: from mechanistic insights to therapeutic opportunities. Clinical and experimental pharmacology & physiology. 2004 Mar:31(3):119-28 [PubMed PMID: 15008953]
Baena-Lopez LA, Wang L, Wendler F. Cellular stress management by caspases. Current opinion in cell biology. 2024 Feb:86():102314. doi: 10.1016/j.ceb.2023.102314. Epub 2024 Jan 11 [PubMed PMID: 38215516]
Level 3 (low-level) evidenceHu SJ, Jiang SS, Zhang J, Luo D, Yu B, Yang LY, Zhong HH, Yang MW, Liu LY, Hong FF, Yang SL. Effects of apoptosis on liver aging. World journal of clinical cases. 2019 Mar 26:7(6):691-704. doi: 10.12998/wjcc.v7.i6.691. Epub [PubMed PMID: 30968034]
Level 3 (low-level) evidenceNguyen TT, Wei S, Nguyen TH, Jo Y, Zhang Y, Park W, Gariani K, Oh CM, Kim HH, Ha KT, Park KS, Park R, Lee IK, Shong M, Houtkooper RH, Ryu D. Mitochondria-associated programmed cell death as a therapeutic target for age-related disease. Experimental & molecular medicine. 2023 Aug:55(8):1595-1619. doi: 10.1038/s12276-023-01046-5. Epub 2023 Aug 23 [PubMed PMID: 37612409]
Kurokawa M, Kornbluth S. Caspases and kinases in a death grip. Cell. 2009 Sep 4:138(5):838-54. doi: 10.1016/j.cell.2009.08.021. Epub [PubMed PMID: 19737514]
Duarte-Olivenza C, Moran G, Hurle JM, Lorda-Diez CI, Montero JA. Lysosomes, caspase-mediated apoptosis, and cytoplasmic activation of P21, but not cell senescence, participate in a redundant fashion in embryonic morphogenetic cell death. Cell death & disease. 2023 Dec 9:14(12):813. doi: 10.1038/s41419-023-06326-6. Epub 2023 Dec 9 [PubMed PMID: 38071330]
Nössing C, Ryan KM. 50 years on and still very much alive: 'Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics'. British journal of cancer. 2023 Feb:128(3):426-431. doi: 10.1038/s41416-022-02020-0. Epub 2022 Nov 11 [PubMed PMID: 36369364]
Yin S, Ji C, Wu P, Jin C, Qian H. Human umbilical cord mesenchymal stem cells and exosomes: bioactive ways of tissue injury repair. American journal of translational research. 2019:11(3):1230-1240 [PubMed PMID: 30972158]
Level 2 (mid-level) evidenceMehrbod P, Ande SR, Alizadeh J, Rahimizadeh S, Shariati A, Malek H, Hashemi M, Glover KKM, Sher AA, Coombs KM, Ghavami S. The roles of apoptosis, autophagy and unfolded protein response in arbovirus, influenza virus, and HIV infections. Virulence. 2019 Dec:10(1):376-413. doi: 10.1080/21505594.2019.1605803. Epub [PubMed PMID: 30966844]
Chaudhry GE, Akim AM, Sung YY, Muhammad TST. Cancer and Apoptosis. Methods in molecular biology (Clifton, N.J.). 2022:2543():191-210. doi: 10.1007/978-1-0716-2553-8_16. Epub [PubMed PMID: 36087269]
Tian X, Srinivasan PR, Tajiknia V, Sanchez Sevilla Uruchurtu AF, Seyhan AA, Carneiro BA, De La Cruz A, Pinho-Schwermann M, George A, Zhao S, Strandberg J, Di Cristofano F, Zhang S, Zhou L, Raufi AG, Navaraj A, Zhang Y, Verovkina N, Ghandali M, Ryspayeva D, El-Deiry WS. Targeting apoptotic pathways for cancer therapy. The Journal of clinical investigation. 2024 Jul 15:134(14):. doi: 10.1172/JCI179570. Epub 2024 Jul 15 [PubMed PMID: 39007268]
Thatte U, Dahanukar S. Apoptosis: clinical relevance and pharmacological manipulation. Drugs. 1997 Oct:54(4):511-32 [PubMed PMID: 9339959]
Croce CM, Tait SWG, Garcia-Sáez AJ, Villunger A, Letai A, Walter HS, Dyer MJS, Green DR, Shi Y, Melino G. What does BCL-2 do? From new molecular insights to the clinical implications. Cell death and differentiation. 2026 Apr:33(4):673-693. doi: 10.1038/s41418-025-01607-3. Epub 2026 Feb 19 [PubMed PMID: 41714805]
Pistritto G, Trisciuoglio D, Ceci C, Garufi A, D'Orazi G. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging. 2016 Apr:8(4):603-19. doi: 10.18632/aging.100934. Epub [PubMed PMID: 27019364]
Lorenz HM, Herrmann M, Winkler T, Gaipl U, Kalden JR. Role of apoptosis in autoimmunity. Apoptosis : an international journal on programmed cell death. 2000 Nov:5(5):443-9 [PubMed PMID: 11256887]
Radi E, Formichi P, Battisti C, Federico A. Apoptosis and oxidative stress in neurodegenerative diseases. Journal of Alzheimer's disease : JAD. 2014:42 Suppl 3():S125-52. doi: 10.3233/JAD-132738. Epub [PubMed PMID: 25056458]
Erekat NS. Apoptosis and its therapeutic implications in neurodegenerative diseases. Clinical anatomy (New York, N.Y.). 2022 Jan:35(1):65-78. doi: 10.1002/ca.23792. Epub 2021 Oct 7 [PubMed PMID: 34558138]
Sharma VK, Singh TG, Singh S, Garg N, Dhiman S. Apoptotic Pathways and Alzheimer's Disease: Probing Therapeutic Potential. Neurochemical research. 2021 Dec:46(12):3103-3122. doi: 10.1007/s11064-021-03418-7. Epub 2021 Aug 12 [PubMed PMID: 34386919]
Wang K, Zhu Q, Liu W, Wang L, Li X, Zhao C, Wu N, Ma C. Mitochondrial apoptosis in response to cardiac ischemia-reperfusion injury. Journal of translational medicine. 2025 Jan 28:23(1):125. doi: 10.1186/s12967-025-06136-8. Epub 2025 Jan 28 [PubMed PMID: 39875870]
Xiang M, Lu Y, Xin L, Gao J, Shang C, Jiang Z, Lin H, Fang X, Qu Y, Wang Y, Shen Z, Zhao M, Cui X. Role of Oxidative Stress in Reperfusion following Myocardial Ischemia and Its Treatments. Oxidative medicine and cellular longevity. 2021:2021():6614009. doi: 10.1155/2021/6614009. Epub 2021 May 18 [PubMed PMID: 34055195]
D'Oria R, Schipani R, Leonardini A, Natalicchio A, Perrini S, Cignarelli A, Laviola L, Giorgino F. The Role of Oxidative Stress in Cardiac Disease: From Physiological Response to Injury Factor. Oxidative medicine and cellular longevity. 2020:2020():5732956. doi: 10.1155/2020/5732956. Epub 2020 May 14 [PubMed PMID: 32509147]
Moyer A, Tanaka K, Cheng EH. Apoptosis in Cancer Biology and Therapy. Annual review of pathology. 2025 Jan:20(1):303-328. doi: 10.1146/annurev-pathmechdis-051222-115023. Epub [PubMed PMID: 39854189]
Diepstraten ST, Anderson MA, Czabotar PE, Lessene G, Strasser A, Kelly GL. The manipulation of apoptosis for cancer therapy using BH3-mimetic drugs. Nature reviews. Cancer. 2022 Jan:22(1):45-64. doi: 10.1038/s41568-021-00407-4. Epub 2021 Oct 18 [PubMed PMID: 34663943]
Vogler M, Braun Y, Smith VM, Westhoff MA, Pereira RS, Pieper NM, Anders M, Callens M, Vervliet T, Abbas M, Macip S, Schmid R, Bultynck G, Dyer MJ. The BCL2 family: from apoptosis mechanisms to new advances in targeted therapy. Signal transduction and targeted therapy. 2025 Mar 21:10(1):91. doi: 10.1038/s41392-025-02176-0. Epub 2025 Mar 21 [PubMed PMID: 40113751]
Level 3 (low-level) evidenceYao X, Lu Y, Shi S, Wang Y. Antibody-drug conjugates (ADCs): "Smart Missiles" targeting cancer therapy. Colloids and surfaces. B, Biointerfaces. 2026 Jan:257():115161. doi: 10.1016/j.colsurfb.2025.115161. Epub 2025 Sep 25 [PubMed PMID: 41015828]
Xu X, Yu T, Wang Z. Combining antibody-drug conjugates with immune checkpoint inhibitors: A new paradigm for breast cancer therapy. Cancer treatment reviews. 2025 Nov:140():103012. doi: 10.1016/j.ctrv.2025.103012. Epub 2025 Aug 11 [PubMed PMID: 40815982]
Khalef L, Lydia R, Filicia K, Moussa B. Cell viability and cytotoxicity assays: Biochemical elements and cellular compartments. Cell biochemistry and function. 2024 Apr:42(3):e4007. doi: 10.1002/cbf.4007. Epub [PubMed PMID: 38593323]
Kockx MM, Muhring J, Knaapen MW, de Meyer GR. RNA synthesis and splicing interferes with DNA in situ end labeling techniques used to detect apoptosis. The American journal of pathology. 1998 Apr:152(4):885-8 [PubMed PMID: 9546348]
Cilloni D, Ferrando A, Frassoni F. Targeting BCL-xL in Myeloid Malignancies: From Inhibitors to PROTAC. Journal of cellular and molecular medicine. 2026 Apr:30(7):e71036. doi: 10.1111/jcmm.71036. Epub [PubMed PMID: 41937490]